Article Text

Abstract
Background: Oxidative stress in RA synovial tissue can cause DNA damage and suppress the DNA mismatch repair (MMR) system in cultured synoviocytes. This mechanism includes two enzyme complexes, hMutSα (hMSH2/hMSH6) and hMutSβ (hMSH2/hMSH3).
Objective: To examine the expression and distribution of MMR enzymes in synovial tissues from patients with arthritis and from normal subjects.
Methods: Synovial tissues from patients with RA, osteoarthritis (OA), or normal subjects were analysed by immunohistochemistry using monoclonal antibodies to hMSH2, hMSH3, and hMSH6. MMR protein expression was evaluated by computer assisted digital image analysis.
Results: hMSH2, hMSH3, and hMSH6 were found in most synovial tissues evaluated, with greater levels in the intimal lining than sublining regions. In RA and OA, sublining perivascular staining for hMSH6 and hMSH3 was also prominent. Significantly higher sublining expression of hMSH2, hMSH3, and hMSH6 was seen in RA and OA than in normal synovium. Double label immunohistochemistry demonstrated that the main cells expressing MMR enzymes were CD68+ and CD68− cells in the intimal lining.
Conclusions: DNA MMR enzyme expression is greatest in the synovial intimal lining layer, where maximal oxidative stress in RA occurs. Although MMR enzyme expression is greater in RA than in normal tissue, this compensatory response cannot overcome the genotoxic environment, and DNA damage accumulates.
- AP, alkaline phosphatase
- DAB, 3, 3′-diaminobenzidine
- hMSH, human MutS homologue
- HPF, high power field
- HRP, horseradish peroxidase
- mAb, monoclonal antibody
- MMR, DNA mismatch repair
- OA, osteoarthritis
- RA, rheumatoid arthritis
- ST, synovial tissue
- DNA damage
- immunohistochemistry
- osteoarthritis
- rheumatoid arthritis
- synovium
Statistics from Altmetric.com
- AP, alkaline phosphatase
- DAB, 3, 3′-diaminobenzidine
- hMSH, human MutS homologue
- HPF, high power field
- HRP, horseradish peroxidase
- mAb, monoclonal antibody
- MMR, DNA mismatch repair
- OA, osteoarthritis
- RA, rheumatoid arthritis
- ST, synovial tissue
Rheumatoid arthritis (RA) is a chronic inflammatory disease characterised by synovial hyperplasia and progressive joint damage.1 Extracellular matrix destruction in RA is mediated, in part, by activated osteoclasts, production of proteolytic enzymes, and tissue invasion by aggressive synoviocytes.2 Although the mechanisms that enhance synoviocyte invasiveness are not fully understood, genetic alterations caused by oxidative stress might contribute. For instance, somatic mutations in genomic microsatellites3 and in genes that can regulate matrix homoeostasis, including p53,4–7 hprt,8,9 WISP-3,9a and h-ras,10 have been found in RA synovium and synoviocytes.
DNA mismatch repair (MMR) mechanisms counteract the genotoxic environment encountered during cell replication. In humans, the principal MMR enzymes include the prokaryotic MutS homologues hMSH2, hMSH3, and hMSH6.11 These proteins form two heterodimers: hMutSα (a heterodimer of hMSH2 and hMSH6) and hMutSβ (a heterodimer of hMSH2 and hMSH3). The former primarily repairs single base mismatches, whereas the latter corrects longer insertion/deletion loops.12 Previous studies have shown that MMR enzymes are expressed in RA synovium and that oxidative stress selectively suppresses MSH6 expression in cultured synoviocytes. However, little is known about the distribution of MMR protein expression in RA compared with osteoarthritis (OA) and normal synovium. Therefore, we performed immunohistochemistry with digital image analysis to characterise the expression of hMSH2, hMSH3, and hMSH6. These studies show that the synovial intimal lining, which is a primary site of synovial mutations and oxidative damage in RA, is also the main location for MMR expression.
MATERIALS AND METHODS
Synovial tissues
Synovial tissue (ST) was obtained at the time of joint replacement for patients with RA and OA and at the time of necropsy for normal subjects without arthritis. The mean (SD) ages of the patients with RA, OA, and normal subjects were 58.5 (4.6), 75 (4.6), and 51 (5.5), respectively. All patients with RA fulfilled the American College of Rheumatology criteria.13 The study protocol was approved by the UCSD Human Research Protection Program. STs were snap frozen in Tissue-Tek OCT compound (Sakura Finetek USA Inc, Torrance, CA, USA) and stored at −80°C until used. Frozen sections (5 μm) were mounted on poly-l-lysine coated Poly-Prep glass slides (Sigma, St Louis, MO, USA) and fixed in acetone (Sigma, St Louis, MO, USA).
Immunohistochemistry
Serial sections were stained with the following mouse monoclonal antibodies (mAbs): hMSH2 (2 μg/ml, Ab-2; Oncogene Research Products, San Diego, CA, USA), hMSH3 (5 μg/ml; Becton Dickinson Biosciences, San Diego, CA, USA), and hMSH6 (5 μg/ml, MSH6/GTBP; Becton Dickinson Biosciences). For control sections, matched concentrations of mouse isotype control Ab (DAKO, Glostrup, Denmark) were used. Immunohistochemical staining was performed as previously described.14 After overnight incubation with the primary mAb, sections were washed, incubated for 30 minutes with biotinylated horse antimouse Ab (Vector Laboratories, Burlingame, CA, USA), washed extensively, and then depleted of endogenous peroxidase. The tissue sections were subsequently stained using the biotin-streptavidin/peroxidase method (Vector Laboratories), and horseradish peroxidase (HRP) activity was detected with hydrogen peroxide and 3, 3′-diaminobenzidine (DAB) (Sigma, St Louis, MO, USA). Slides were counterstained with Mayer’s haematoxylin (DAKO).
For double label immunohistochemistry studies, the ST sections were blocked with 1% bovine serum albumin, 5% horse serum and 10% normal human serum, and first stained with the anti-MSH Ab as described above, except that HRP activity was detected with 3-amino-9-ethylcarbazole. The second primary Ab was applied overnight at 4°C. To detect macrophages, goat anti-CD68 (Santa Cruz Biotechnology Inc, Santa Cruz, CA, USA) was used after blocking with 5% swine serum. Alkaline phosphatase (AP) conjugated swine antigoat Ab (Southern Biotechnology Associates Inc, Birmingham, AL, USA) served as the secondary Ab. To detect T cells, rabbit anti-CD3 (Zymed Laboratories Inc, South San Francisco, CA, USA) was used after blocking with 5% goat serum. AP conjugated goat antirabbit Ab (Biomeda, Foster City, CA) served as the secondary Ab. AP activity was detected with levamisole and Vector Blue (Vector Laboratories).
Digital image analysis
Sections were analysed in a random order by digital image analysis.15 Five to six high power fields (HPF) images (×400 magnification) for each synovial section were acquired on a Nikon Eclipse E800 microscope and captured using a charged coupled device Olympus MicroFire digital camera and MicroFire 2.1 software (Olympus America, Melville, NY, USA). The image acquisition process was standardised by calibrating the microscope and the camera for each MSH protein. Each HPF evaluated contained clearly identifiable synovial intimal lining and sublining regions, and was analysed with Image-ProPlus image analysis program (version 4.5 for Windows; Media Cybernetics Inc, Silver Spring, MD, USA). For each MSH protein, two colour discrimination thresholds were adjusted by the colour cube based segmentation to create two binary overlay masks. The first binary mask was selected to cover the positively stained area (DAB), and the second one covered the total region of tissue (DAB and haematoxylin). The percentage of pixels above the designated colour threshold in the region of interest (intimal lining or sublining) was determined for each HPF.
Statistical analysis
Analysis of variance was used to test for differences between the groups (Tukey–Kramer honestly significant difference). For the data within each study group Wilcoxon’s non-parametric test was used. A value of p<0.01 was considered significant.
RESULTS AND DISCUSSION
To evaluate the distribution and relative expression of DNA MMR enzymes, immunohistochemistry was performed on synovium from patients with RA and OA and from normal subjects. Paraffin embedded sections of normal colon and cytospun samples of formalin fixed colorectal cancer cell lines (HCT116 and SW480) served as positive controls (data not shown). Expression of all three MMR enzymes (hMSH2, hMSH3, and hMSH6) was demonstrated in virtually all STs evaluated (n = 6, 5, and 4 for RA, OA, and normal subjects, respectively) (see fig 1 for representative examples of each). Image analysis demonstrated significantly greater expression of the three enzymes in the synovial intimal lining than in the sublining layer (p<0.01) (table 1, fig 2). In addition to high expression in the intimal lining, hMSH6, and hMSH3 also demonstrated prominent staining in the perivascular regions in both RA and OA ST (fig 1). The expression pattern for hMSH2 in the sublining layer was similar to a previous report for RA synovium16 (fig 1).
Statistical analysis of MMR enzyme expression in synovium

Distribution of DNA MMR enzymes in synovial tissue. Sections from RA (n = 6), OA (n = 5), and normal (n = 4) synovia were incubated with mAb to hMSH2, hMSH3, or hMSH6. The proteins were detected using the immunoperoxidase technique and DAB (brown colour). The slides were counterstained with haematoxylin (blue). Representative sections are shown. Images were obtained at ×200 magnification, and each insert obtained at ×400 magnification.
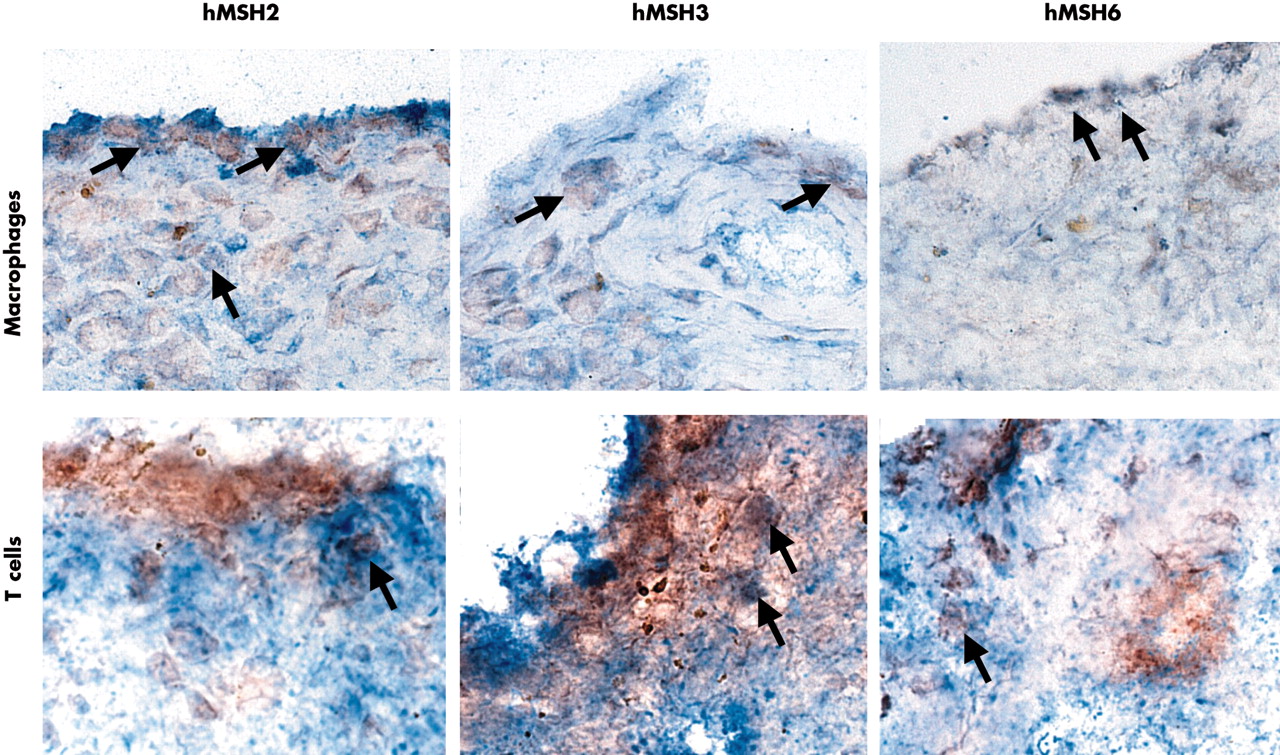
Double label immunohistochemistry for MMR enzymes and cell surface markers. Representative field in an RA synovium are shown demonstrating the expression of hMSH2, hMSH3, and hMSH6 in CD3+ T cells and CD68+ macrophages. Examples of double positive cells are indicated by arrows. The most abundant staining for all three MMR proteins (red, peroxidase) was seen in intimal lining CD68+ (blue, AP) and CD68− cells (macrophage-like synoviocytes and fibroblast-like synoviocytes, respectively). Only rare T cells (blue, AP) in the sublining were positive (an example is shown in the figure).
When DNA MMR enzyme expression in RA was compared with OA, hMSH3 and hMSH6 levels as determined by the percentage of the region containing the protein were modestly higher in RA (p<0.01). However, hMSH2 expression in RA was similar to OA (table 1, figs 1 and 2). The data on hMSH6 expression compare with the results of our recent study of synovial microsatellite instability, in which hMSH6 expression was decreased in RA compared with OA by western blot analysis.3 This apparent difference is probably due to the techniques employed. Western blot provides an accurate representation of normalised protein expression but lacks the spatial resolution of immunohistochemistry. Despite this limitation, the immunostaining can identify the primary sites of protein expression and can give insights into the percentage of the tissue that expresses the protein of interest. Therefore, these two techniques provide complementary information.
To determine which cells in the synovium express the MMR proteins, double label immunohistochemistry was performed. As shown in fig 2, CD68+ and CD68− cells in the intimal lining expressed hMSH2, hMSH3, and hMSH6 most abundantly. Occasional sublining CD68 macrophages, but only rare CD3 T cells, were positive for MMR proteins.
In addition to synovium from patients with arthritis, MMR distribution was also evaluated in normal synovium. The synovial intimal lining was, again, the primary site of MMR enzyme expression, although the percentage of synovium that stained was significantly lower in normal tissue than in arthritis tissue (table 1, fig 3). These data indicate that MMR proteins are normally expressed in ST, especially in the intimal lining. Although the function of MMR enzymes was not assessed, previous studies demonstrate that the level of MMR enzyme expression correlates with MMR activity.17 It is likely, therefore, that DNA repair activity is markedly higher in synovium from arthritis patients than in normal joint tissue.

{kind=link}
{kind=link}
{kind=link}
Digital image analysis of DNA MMR enzyme expression in ST. Intimal lining and sublining regions were defined in five to six HPF images (×400 magnification) from each ST. The percentage positive area was determined by dividing the number of pixels above a colour threshold by the total number of pixels within the selected area. The data are presented as mean (SEM). *p<0.003 when comparing lining and sublining regions in RA, OA, or normal synovium; **p<0.01 when comparing lining or sublining regions of RA with OA; # p<0.004 when comparing RA or OA with normal synovium.
The increase in MMR expression in arthritis may relate to oxidative stress encountered during inflammatory responses. Proinflammatory cytokines do not alter MMR enzyme levels in synoviocytes, whereas exposure to reactive oxygen and nitrogen has complex effects that depend on the specific MSH protein examined.3 We have previously demonstrated that somatic mutations occur in certain key genes like p53 in RA but not OA synovium, especially in the intimal lining.18 Furthermore, oxidative stress and DNA damage are significantly greater in RA synovium than in OA.19 Possibly, the compensatory increase of MMR expression seen in RA and OA synovium reaches maximum levels, thereby overwhelming the repair mechanisms in inflammatory synovitis. Under these circumstances, a burden of mutations could accumulate over time in RA even though MMR enzyme expression may be greater than in normal tissue and similar to that in OA synovium. The relative lack of MMR expression of T cells is consistent with the observation that certain somatic mutations also occur in articular T cells.8
Although the functional sequelae of mutations in rheumatoid synovium remain uncertain, the possibility that DNA damage overwhelms repair mechanisms suggests potential therapeutic interventions that might be beneficial in RA. For instance, efforts to either decrease production of reactive oxygen or enhance MMR production might potentially limit the accumulations of mutations. By maintaining DNA integrity, genotoxic events that possibly alter the natural history of disease might be prevented.
Acknowledgments
This study was supported by National Institutes of Health (grants AR45347 and AR47825) and the UCSD Rheumatic Diseases Core Center (AR47360).