Article Text

Abstract
Objectives Nerve growth factor (NGF) is a promising analgesic target, particularly in osteoarthritis (OA) where existing therapies are inadequate. We hypothesised that pain responses to NGF are increased in OA joints. Here, NGF-evoked pain behaviour was compared in two rodent models of OA, and possible mechanisms underlying altered pain responses were examined.
Methods OA was induced in rat knees by meniscal transection (MNX) or intra-articular monosodium iodoacetate injection (MIA). Once OA pathology was fully established (day 20), we assessed pain behaviour (hindlimb weight-bearing asymmetry and hindpaw mechanical withdrawal thresholds) evoked by intra-articular injection of NGF (10 µg). Possible mechanisms underlying alterations in NGF-induced pain behaviour were explored using indomethacin pretreatment, histopathological evaluation of synovitis, and rtPCR for NGF receptor (tropomyosin receptor kinase (Trk)-A) expression in dorsal root ganglia (DRG).
Results Both the MIA and MNX models of OA displayed reduced ipsilateral weight bearing and hindpaw mechanical withdrawal thresholds, mild synovitis and increased TrkA expression in DRG. NGF injection into OA knees produced a prolonged augmentation of weight-bearing asymmetry, compared to NGF injection in non-osteoarthritic knees. However, hindpaw mechanical withdrawal thresholds were not further decreased by NGF. Pretreatment with indomethacin attenuated NGF-facilitated weight-bearing asymmetry and reversed OA-induced ipsilateral TrkA mRNA up-regulation.
Conclusions OA knees were more sensitive to NGF-induced pain behaviour compared to non-osteoarthritic knees. Cyclo-oxygenase products may contribute to increased TrkA expression during OA development, and the subsequent increased NGF sensitivity. Treatments that reduce sensitivity to NGF have potential to improve OA pain.
- Knee Osteoarthritis
- Corticosteroids
- Synovitis
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/
Statistics from Altmetric.com
Background
Pain is the predominant feature of osteoarthritis (OA),1 and most people with OA continue to experience pain despite analgesic use. OA pain may be augmented through multiple mechanisms, both within the joint and through sensitisation of pain processing pathways. Synovitis and joint damage are closely associated with OA pain,2–4 and cyclo-oxygenase inhibitors and intra-articular glucocorticosteroid injections can reduce both inflammation and pain in man and in rat OA models.2 ,5–7 Inflammation contributes to pain flares in OA, exacerbates articular damage, and may initiate sustained sensitisation that subsequently augments chronic pain. Unfortunately, the benefits of anti-inflammatory treatments in OA are often incomplete, not sustained, and may be associated with significant adverse events. There is an urgent need for new analgesic agents for people with OA.
Nerve growth factor (NGF) plays a key role in many persistent pain states, notably those associated with inflammation.8–13 The β subunit of NGF binds tropomyosin receptor kinase-A (TrkA) and p75 neurotrophin receptors on sensory nerve terminals.14 ,15 Exogenous administration of small doses of NGF to adult animals and humans can produce pain and hyperalgesia.16–18 NGF is up-regulated in human19 and rodent20 osteoarthritic joints and is associated with inflammatory cell infiltration of the synovium9 ,21–23 and subchondral bone.24 ,25 Clinical trials with NGF-blocking antibodies indicate NGF makes an important contribution to OA pain.26 Indeed, the effects in OA appear superior to those in other chronic pain states, such as neuropathic pain,27 suggesting a particular contribution of NGF to OA pain. We hypothesised that the importance of NGF for OA pain may be due, in part, to increased sensitivity of OA joints to its pain-augmenting actions and that inflammation may contribute to this increased sensitivity to NGF.
We aimed to determine the effects of intra-articular NGF injection in normal and osteoarthritic rat knees, and to explore possible mechanisms underlying any altered sensitivity to NGF by measuring mRNA expression of the high affinity NGF receptor, TrkA, and by inhibiting inflammation during OA pathogenesis.
Materials and methods
Animals models of OA
In vivo experiments performed on 7-week-old male Sprague–Dawley rats (n=8 per group, weighing 200–220 g; Charles River, Kent, UK) were carried out in accordance with UK Home Office regulations and follow the guidelines of the International Association for the Study of Pain. Rats were housed under standard conditions under a 12 h light/dark cycle, with unlimited access to food and water. Rats were anaesthetised with isoflurane (2% in O2) prior to surgery and intra-articular injections.2 All outcome measurements were made by an observer blinded to treatment. Non-osteoarthritic rats were used as controls. Experimental designs and group sizes are described in online supplementary figures S1 and S2.
Induction of OA
OA was induced on day 0 in the left knee by either medial meniscal transection (MNX)2 or by a single intra-articular injection of monosodium iodoacetate (MIA; 1 mg/50 µL sterile 0.9% normal saline, pH 7.4). MIA dose was based on previous studies.28 ,29 Controls received intra-articular saline injection. We previously demonstrated that sham-operated controls do not display pain behaviour, inflammation or OA structural changes beyond 7 days after surgery.30 ,31
Pharmacological interventions
Intra-articular NGF injection
Rats were given a single 50 µL intra-articular injection into their left knees of NGF in sterile 0.9% normal saline, pH 7.4, or saline control. Initial dose–response studies in normal rats used 10 µg, 1 µg or 100 ng of NGF. Subsequent experiments compared the effects of NGF (10 µg/50 µL) or saline control injected into the left knee of non-osteoarthritic rats and rats 20 days after OA induction. By day 20, OA pathology and pain behaviour are evident in both the MIA and MNX models of OA.2 ,32
Systemic indomethacin administration
To investigate the contribution of OA-associated synovitis to altered NGF-induced pain responses, rats with MIA-induced OA received daily oral gavage with 0.5 mL of either 2 mg/kg indomethacin or vehicle (sterile 0.9% normal saline, pH 7.4) from 1 day before OA induction continuing until day 18. Indomethacin treatment ceased 2 days before NGF injection in order to avoid any direct effects of indomethacin on pain behaviour. This dose of indomethacin showed attenuated pain behaviour and inflammation in earlier reports.2 ,33
Pain behaviour and joint swelling
Pain behaviour was measured as hindlimb weight-bearing asymmetry and as distal allodynia to punctate stimulation of the hindpaw. The difference in weight bearing (g) between the ipsilateral treated limb and the contralateral control limb was measured using an incapacitance meter (Linton Instruments, Norfolk, UK) as the average of five readings.2 Paw withdrawal thresholds were measured using a series of von Frey monofilaments (Semmes-Weinstein monofilaments with bending forces of 1, 1.4, 2, 4, 6, 8, 10 and 15 g) applied in ascending order of bending force to the plantar surface.32 The paw withdrawal threshold was the lowest weight of monofilament that elicited a withdrawal reflex.
Joint swelling was measured with digital electronic callipers (Mitutoyo, Andover, UK) as the difference in knee diameter (millimeters) between the ipsilateral and contralateral knees.
Tissue processing
At the termination of each experiment rats were killed by asphyxiation in carbon dioxide, and synovia with patellae from the right and left knees were immediately harvested, embedded in OCT and snap frozen over melting isopentane. Ipsilateral and contralateral dorsal root ganglia (DRG) from lumbar regions 1, 2, 3, 4 and 5 were frozen on dry ice. Tibiofemoral joints were fixed for 48 h in neutral buffered formalin (containing 4% paraformaldehyde), then decalcified in 10% ethylenediaminetetraacetic acid in 10 mM Tris buffer (pH 6.95) for 4 weeks at room temperature. Coronal sections of trimmed joint tissues were mounted in paraffin wax.
Joint pathology
Sections for histology were cut at 5 μm and visualised using a 20× objective lens unless otherwise indicated. Synovial lining thickness and cellularity were scored using H&E-stained sections.30 Synovial macrophage infiltration was identified by immunoreactivity for the monoclonal antibody clone ED1 directed to CD68,34 and developed with diaminobenzidine using the glucose oxidase/nickel-enhanced method.35 The macrophage fractional area was the percentage of synovial section area positive for CD68, derived using four fields per section and one section per case.35 ,36 Mid-coronal knee sections were identified by the presence of the cruciate ligament insertions, and stained with Safranin-O. Osteochondral integrity was measured as the number of channels present in the articular cartilage across the entire medial tibial plateau of one section.2 Chondropathy and osteophytosis were evaluated as previously described using a 4× objective lens.2 ,31 All image analyses were performed by an observer blinded to experimental details using a Zeiss Axioskop 50 microscope (Carl Zeiss, Welwyn Garden City, UK) and a KS300 image analysis system (Image Associates, Thame, UK).
RNA extraction and cDNA synthesis
DRG were pooled to produce four separate samples from each rat, derived from DRG innervating (L3–5) or rostral to (L1–2) the ipsilateral or contralateral knee. Spinal cords (L1–6) were used as control tissues due to their low levels of TrkA expression.37 Tissues were homogenised and RNA purified using the Nucleospin RNA II Kit (Macherey-Nagel). For cDNA synthesis, 200 ng of total RNA was reverse transcribed using AffinityScript reverse transcriptase (Agilent Technologies) in a total reaction volume of 24 µL. Reactions were incubated for 5 min at 25°C, 1 h at 50°C and terminated by incubation at 70°C for 15 min.
TaqMan quantitative real time PCR
Gene expression was quantified utilising the relative standard curve method based on TaqMan quantitative real time PCR (qRT-PCR). Primers and probes for β-actin were obtained from published work,29 and those for TrkA were designed using Primer Express software and synthesised by MWG Biotech (Germany). Data are expressed as a ratio of gene expression levels with reference to β-actin. TrkA forward primer: 5-GGGAACCTGACGGAGCTCTAT-3, reverse primer: 5-ACAAAGCGGAGGCCACTCT-3, TaqMan probe: 5-CCAGCGGGATCTGCAACGCCT-3. β-Actin forward primer: 5-AGGCCATGTACGTAGCCATCCA-3, reverse primer: 5-TCTCCGGAGTCCATCACAATG-3, TaqMan probe: 5-TGTCCCTGTATGCCTCTGGTCGTACCAC-3.
Statistical analysis
Data were analysed using the Statistical Package for the Social Sciences V.16 (SPSS, Chicago, Illinois, USA) and graphically presented using Prism V.4 (GraphPad, San Diego, California, USA). Parametric data (macrophage fractional areas (logarithmically transformed), incapacitance and knee diameter) were analysed using one way analysis of variance (ANOVA). Univariate comparisons were made against controls using the Student t test. Non-parametric data (joint pathology) were analysed using the Kruskal–Wallis test followed by the Mann–Whitney test with Bonferroni correction. Numerical data are quoted as mean (95% CI) in the text, and, for clarity, graphically as mean±SEM unless otherwise stated. A two-tailed p<0.05 was taken to indicate statistical significance.
Reagents
Biotinylated rat-adsorbed horse anti-mouse antibody and avidin–biotin complexes (ABC Kits) were from Vector Laboratories (Peterborough, UK). Monoclonal antibody to macrophages (clone ED1) was from Serotec (Oxford, UK). NGFβ (N2513), indomethacin and all other chemicals were obtained from Sigma-Aldrich (Poole, UK).
Results
Intra-articular injection of NGF induces pain behaviour and synovitis
Intra-articular injection of NGF produced, at 24 h, dose dependent increases in pain behaviour (weight-bearing asymmetry and differences in paw withdrawal thresholds; figure 1A,C), joint swelling (figure 1E) and synovial macrophage infiltration (figure 1F) in non-osteoarthritic rats. Responses returned to control values within 3–5 days (figure 1B,D–F). On the basis of these experiments, the 10 µg dose of NGF was selected for use in the OA pain models.

Nerve growth factor (NGF)-induced pain behaviour and inflammation in non-arthritic rat knee joints. Rat knees (n=8 rats/group) were injected with NGF 100 ng (•), 1 µg (▪), 10 µg (▴) or saline (◊) on day 0. There was a dose dependent increase in pain behaviour, measured as the difference in weight-bearing distribution between the hindlimbs (A and B) and paw withdrawal threshold to von Frey hair stimulation (C and D), joint swelling (E) and synovial macrophage infiltration (F) 24 h after NGF injection. Pain behaviour and joint swelling had normalised to saline control levels at 24 h after intra-articular injection of 100 ng and 1 µg NGF (A, C and E). Pain behaviour and joint swelling were increased 1 h after intra-articular injection of 10 µg NGF in non-osteoarthritic rat knees, reached a maximum at 3 h and normalised to saline controls by days 3–5 (A–E). No statistically significant difference in pain behaviour measured as weight-bearing asymmetry (B) and joint swelling (E) was seen beyond day 1. Synovitis had also normalised to saline control levels by day 7 (F). +++p<0.001 versus naive control. *p<0.05, **p<0.01, ***p<0.001 versus saline control.
Effects of OA on NGF-induced pain behaviour
OA pathology was fully developed 20 days after MNX or intra-articular MIA injection, and persisted through day 35 (table 1). OA characteristics in both models were: increased pain behaviour and joint diameter (figure 2), chondropathy, osteophytosis and mild synovitis including macrophage infiltration of synovium (see online supplementary table 1 and figure S3).
Structural changes in the meniscal transection (MNX) and monosodium iodoacetate (MIA) animal models of osteoarthritis at day 20 and 35 after induction

Augmented nerve growth factor (NGF)-induced weight-bearing asymmetry, but not paw withdrawal thresholds or joint swelling, in rat models of osteoarthritis (OA). NGF-induced pain behaviour and inflammation in meniscal transection (MNX) (A, C and E) and monosodium iodoacetate (MIA) (B, D and F) models of OA. Pain behaviour, measured as increased hindlimb weight-bearing asymmetry (A and B) and reduced paw withdrawal thresholds to punctuate mechanical stimulation (C and D), was increased 20 days after the induction of OA in rat knees by either MNX surgery or intra-articular injection of MIA (1 mg/50 µL). In non-osteoarthritic (non-OA) rats, weight-bearing asymmetry increased 2 h following intra-articular injection of 10 µg NGF (▵), and normalised to non-OA (saline-injected) control rats (•) by day 23 (3 days after NGF injection). In MNX or MIA induced OA, weight-bearing asymmetry was further increased 2 h following NGF injection (MNX+NGF (▪), MIA+NGF (⋄)) compared with saline-injected MNX (○) or MIA (▾) rats. Augmented weight-bearing asymmetry after NGF injection in osteoarthritic rats was greater than in non-OA rats, and persisted through day 35. Pain behaviour in NGF-injected non-OA rats had normalised to controls by day 25. Intra-articular injection of NGF reduced paw withdrawal thresholds and increased joint swelling up to 8 h after the injection in the MNX and MIA models of OA but did not further affect paw withdrawal thresholds (C and D) or joint swelling (E and F) in these models of OA. *p<0.05, **p<0.01, ***p<0.001, OA versus non-OA (saline-injected) controls. +p<0.05, +p<0.01, +++p<0.001, OA+NGF rats versus OA (MNX or MIA) rats. #p<0.05, ##p<0.01, ###p<0.001, OA+NGF rats versus non-OA (saline-injected) controls. Vertical broken lines indicate time of NGF injection. n=8 rats/group.
NGF-induced weight-bearing asymmetry was both increased and sustained in osteoarthritic knees compared to NGF effects in non-osteoarthritic knees. Weight-bearing asymmetry increased 2 h after NGF injection into osteoarthritic knees (MNX+NGF: 60.9 (26.8 to 95.0) g, MIA+NGF: 92.3 (61.6 to 123.0) g) compared to saline-injected osteoarthritic rats (MNX: 21.1 (15.6 to 26.7) g, p<0.05; MIA: 21.5 (6.7 to 36.2) g, p<0.001). Increased weight-bearing asymmetry was maintained 7 days after NGF injection into osteoarthritic knees in both the MNX (58.0 (40.0 to 75.9) g) and MIA (42.4 (24.3 to 60.5) g) OA models compared with saline-injected osteoarthritic rats (MNX: 18.8 (12.8 to 24.8) g, p<0.001; MIA: 13.6 (7.6 to 19.6) g, p<0.01). NGF-induced weight-bearing asymmetry in non-osteoarthritic rats had normalised to saline-injected controls by 1 week after the NGF injection, whereas pain behaviour remained augmented in NGF-injected osteoarthritic rats (figure 2A,B).
NGF injection did not further reduce paw withdrawal thresholds (figure 2C,D) or further increase joint swelling (figure 2E,F) in MIA and MNX models of OA, compared to saline-injected osteoarthritic rats. Synovial cellularity and macrophage infiltration 35 days after OA induction did not differ significantly between knees that received NGF or saline injections on day 20 (see online supplementary figure S4).
Effects of indomethacin pretreatment on augmented NGF-induced weight-bearing asymmetry in MIA-induced OA
Weight-bearing asymmetry was reduced during the period of oral treatment with indomethacin (2 mg/kg once daily) from OA induction until day 18 compared to saline-treated osteoarthritic rats (figure 3A). After a 2-day washout (day 20), weight-bearing asymmetry did not differ significantly between indomethacin- and saline-pretreated rats. Indomethacin pretreatment did, however, inhibit the subsequent augmentation by NGF injection of weight-bearing asymmetry in the osteoarthritic rats (40.8 (35.9 to 45.7) compared to osteoarthritic rats which had received saline pretreatment prior to injection of NGF (67.1 (57.7 to 76.6); p<0.001) (figure 3A). Pretreatment with indomethacin had no significant effects on OA-associated synovitis measured before NGF injection (figure 3C,D). NGF injection on day 20 was followed on day 27 by increased synovitis compared with saline-injected osteoarthritic controls (figure 3E,F). Pretreatment with indomethacin did not reduce NGF-induced joint swelling (figure 3B), but reduced synovial macrophage infiltration and synovial lining grade measured 7 days after NGF injection into osteoarthritic knees (figures 3 and 4E,F).

Pretreatment with indomethacin in rats with monosodium iodoacetate (MIA)-induced osteoarthritis (OA) reduced the enhanced and sustained nerve growth factor (NGF)-induced pain behaviour and synovitis, but had no significant effects on NGF-induced joint swelling. In n=8 rats/group, MIA (1 mg/50 µL) or saline (Sal) control was injected in left knee joints on day 0, followed by injection of either NGF (10 µg/50 µL) or saline on day 20 (dotted line) (MIA+saline (•), saline+NGF (▪)). Pretreatment with indomethacin (Indo; 2 mg/kg, orally, daily) or saline control (vehicle (Veh)) was administered from before induction of OA to day 18. Pain behaviour measured as the difference in hindlimb weight bearing (A) was increased following induction of OA. Injection of NGF in non-osteoarthritic (saline-injected) rats produced a short-lived pain response. Injection of NGF in OA rats produced sustained and enhanced pain response (▵) that was reduced following pretreatment with indomethacin (□). Pretreatment with indomethacin had no effect on OA or NGF-induced joint swelling (B). Increased synovitis as measured by synovial macrophage infiltration (C and E) and synovial lining thickness/cellularity (D and F) was observed in osteoarthritic rats compared with non-arthritic controls at days 20 (C and D) and 27 (E and F) but was not significantly reduced following treatment with indomethacin on day 20 (C and D), before NGF injection. NGF injection on day 20 was followed on day 27 by significantly increased synovitis compared with saline-injected osteoarthritic controls. However, pretreatment with indomethacin significantly reduced the enhanced synovitis following NGF injection in rats with MIA-induced OA compared with vehicle pretreated OA rats injected with NGF (day 27; E and F). *p<0.05, **p<0.01, ***p<0.001 versus non-osteoarthritic non-injected rats (control). +p<0.05, ++p<0.01, +++p<0.01, vehicle-treated NGF-injected MIA (MIA+Veh+NGF) rats versus indomethacin-treated NGF-injected MIA (MIA+Indo+NGF) rats. #p<0.05, ##p<0.01, ###p<0.001, MIA+Veh+NGF treated rats versus non-osteoarthritic (saline-injected) NGF-injected (saline+NGF) rats. xp<0.05, xxp<0.01, xxxp<0.001, osteoarthritic saline-injected (MIA+saline) rats versus non-osteoarthritic NGF-injected (saline+NGF) rats. $p<0.05, $$p<0.01, $$$p<0.001, osteoarthritic saline-injected (MIA+saline) rats versus vehicle-treated NGF-injected MIA (MIA+Veh+NGF) rats.
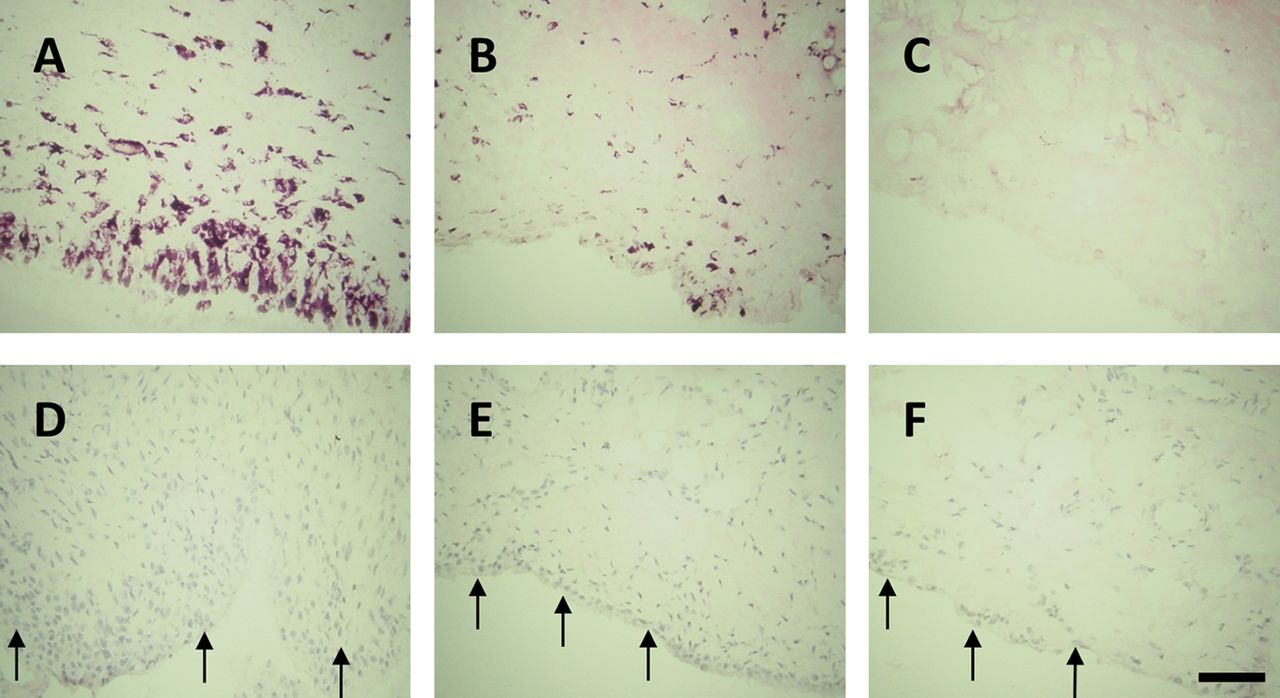
Effect of pretreatment with indomethacin on nerve growth factor (NGF)-induced synovitis. Synovitis 27 days after induction of osteoarthritis (OA) by intra-articular injection of monosodium iodoacetate was measured as macrophage infiltration (delineated by immunoreactivity for CD68; A–C) and synovial lining thickness/cellularity (arrows; D–F). Synovitis was enhanced in NGF-injected OA rats (A and D) at day 27. Pretreatment with indomethacin reduced the enhanced synovitis (synovial macrophage infiltration (B) and synovial lining thickness/cellularity (E)) following NGF injection but not to non-osteoarthritic non-injected control levels (C and F, respectively). Bar=100 µm. n=8 rats/group.
Increased TrkA expression in lumbar DRG from osteoarthritic rats and effects of indomethacin treatment
At 20 and 35 days after OA induction, TrkA expression was increased in the ipsilateral DRG from lumbar regions (L3, 4 and 5) innervating the osteoarthritic knee joint, in both MNX and MIA models of OA compared with non-osteoarthritic control rats (figure 5). TrkA expression was also increased in the contralateral L3, 4 and 5 DRG, and bilaterally in L1 and 2 DRG which do not normally innervate the knee (figure S5). TrkA expression in osteoarthritic rats was reduced following indomethacin treatment (figure 5). Reductions in TrkA expression following pretreatment with indomethacin up to day 18 were statistically significant in L3, 4 and 5 DRG ipsilateral to the osteoarthritic knee 20 days after OA induction (figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Osteoarthritis (OA) induced increases in ipsilateral TrkA expression were reduced by pretreatment with indomethacin. Ipsilateral (A and C) and contralateral (B and D) lumbar (L) 3, 4 and 5 dorsal root ganglia (DRG) of n=8 rats/group innervating the knee joint show a bilateral increase in TrkA mRNA at 20 (A and B) and 35 (C and D) days after induction of OA by either transection of the meniscus (MNX) or intra-articular injection of monosodium iodoacetate (MIA), compared with non-osteoarthritic control rats that received intra-articular saline injection on day 0 (Saline). These effects of indomethacin were statistical significant for ipsilateral (E) but not contralateral (F) L3, 4 and 5 DRG 20 days after OA induction by intra-articular MIA injection. Indomethacin (Indo) (2 mg/kg, orally, daily) or saline control (vehicle (Veh)) was administered from before induction of OA to day 18, followed by a 2-day washout period until sacrifice on day 20. Spinal cord tissues (L1–6) were used as controls. TrkA mRNA is normalised to β-actin mRNA: *p<0.05, **p<0.01, ***p<0.001 versus non-osteoarthritic (saline-injected) rats. +p<0.05 versus vehicle treated MIA rats.
Discussion
We demonstrate that pain behavioural responses to NGF, measured by weight-bearing asymmetry, are increased in OA knees in two rat models. This augmented response to NGF was associated with increased TrkA expression in lumbar DRG and dependent on mechanisms during OA development that can be inhibited by prior treatment with the cyclo-oxygenase inhibitor indomethacin. Increased sensitivity to NGF may, in part, explain the particularly encouraging analgesic effects observed in clinical trials with NGF inhibitors in OA. Reducing sensitivity to NGF has potential to reduce OA pain.
Injection of NGF into non-osteoarthritic rat knees was associated with increased weight-bearing asymmetry and reduced hindpaw withdrawal thresholds to von Frey hair stimulation from 2 h after injection, with normalisation by day 5. Reduced hindpaw withdrawal thresholds may indicate central sensitisation (allodynia at a site distal to the injected joint), whereas the mechanisms underlying weight-bearing asymmetry may include both direct nociception and central sensitisation.18 ,38 Comparable findings in man demonstrate that intramuscular or subcutaneous NGF injection induces pain and both peripheral and central sensitisation. NGF in the knee therefore may have similar effects on pain processing as does NGF in other tissues.
The increased weight-bearing asymmetry observed after intra-articular NGF injection was augmented in osteoarthritic rats compared to non-osteoarthritic rats, with respect to both the magnitude and duration of the response. By contrast, the increase in distal allodynia following intra-articular NGF injection was similar in osteoarthritic and non-osteoarthritic rats. This indicates differential contributions of NGF, and the potential mechanisms underlying weight-bearing asymmetry and distal allodynia may be differentially moderated in OA. TrkA expression was increased in DRG by 20 days after OA induction (the time chosen for testing sensitivity to NGF), and this increased TrkA expression persisted through day 35. Increased TrkA expression has similarly been associated with other rodent models of chronic pain behaviour.39 Increased TrkA expression by sensory nerves innervating the osteoarthritic knee may contribute to the augmented weight-bearing asymmetry following intra-articular injection of NGF.
Although commonly thought of as a degenerative disease of articular cartilage, OA in man is associated with synovitis, characterised by increased synovial cellularity and macrophage infiltration.3 ,4 MIA and MNX OA models similarly displayed mild synovial macrophage infiltration. Cyclo-oxygenase inhibitors such as indomethacin reduce OA pain in man, and, as in previous studies,2 ,40 we found that pain behaviour was reduced during indomethacin treatment in rats with MIA-induced OA. Following discontinuation of indomethacin, pain behaviour returned towards that in saline-treated osteoarthritic controls, consistent with a need for continuing treatment to maintain analgesic efficacy, which, in man, is associated with important risk of gastrointestinal and cardiovascular adverse events.
In addition to these well-recognised effects of cyclo-oxygenase inhibition in OA, pretreatment with indomethacin also reduced NGF-facilitated weight-bearing asymmetry. This may indicate that the enhanced sensitivity to NGF seen at day 20 in the OA models is, at least in part, dependant on cyclo-oxygenase products. Indomethacin pretreatment also inhibited the upregulation of TrkA mRNA in DRG ipsilateral to the osteoarthritic knee, further supporting the hypothesis that increased sensitivity to NGF may be due to increased TrkA expression, which in part may depend on cyclo-oxygenase products.
Indomethacin pretreatment may reduce sensitivity to NGF through several mechanisms, including anti-inflammatory effects in the joint, reduced afferent barrage to the spinal cord, and direct effects on glial cells that contribute to sensitisation in rat OA models.29 NGF-induced inflammation in non-articular tissues involves mast cell degranulation and facilitated neuropeptide release from peripheral nerves.16 ,41 ,42 However, complete suppression of synovitis does not appear to be necessary to prevent increased sensitivity to NGF in OA knees, because synovitis scores and synovial macrophage infiltration remained significantly increased in OA knees at the time of NGF injection even after indomethacin pretreatment, and indomethacin did not significantly reduce joint swelling in the MIA model of OA. The single NGF injection augmented synovitis in OA knees, and this may have contributed to increased weight-bearing asymmetry as both returned to levels observed in non-NGF injected OA knees by day 35. Indomethacin pretreatment reduced the augmented synovitis observed after NGF injection in OA knees, and this may have contributed to reductions in weight-bearing asymmetry.
Experimental studies in animal models have limitations. Insufficient is known about human OA to be certain that key pathological processes are replicated in rodents. However, we have observed similar effects of NGF in both MIA and MNX models of OA, suggesting findings that may be generalisable to OA itself, rather than being dependent on its mode of induction. However, further research is required to determine whether similar mechanisms may operate in human OA, or whether inflammation, cyclo-oxygenases or other mechanisms may also contribute to pain in other OA models. Further research would also be required to determine whether the observed changes in TrkA expression were the causes of altered sensitivity to NGF, and whether contralateral and rostral increases in TrkA expression may parallel the widespread alterations in pain processing observed in patients with OA,43 or whether increased TrkA expression at L1–2 represents rare afferents originating from the knee joint. Apparent differences in the effects of OA on sensitivity to NGF detected by weight-bearing asymmetry and von Frey hair testing may be due to their non-linear measurement properties and further work would be required to confirm our suggestion that NGF-induced allodynia is not altered in models of OA. Despite these reservations, however, our experiments provide evidence that sensitivity to NGF is augmented in OA knees.
NGF blockade is efficacious for OA pain in man.26 However, significant adverse effects on joint integrity44 have raised the possibility that endogenous NGF may play important protective roles in some circumstances. Better understanding of the specific pathways through which NGF augments OA pain, including interactions with cyclo-oxygenases, may identify targets downstream of NGF suitable for novel analgesic development. Our data indicate that synovitis and increased TrkA expression may contribute to the development of NGF sensitivity in OA. Reducing OA-augmented sensitivity to NGF has potential to improve OA pain while retaining the normal homeostatic actions of NGF.
Acknowledgments
The authors would like to thank Paul Millns for his technical assistance with tissue (dorsal root ganglia) collection.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
Footnotes
-
Handling editor Tore K Kvien
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.