Article Text

Abstract
Cellular senescence is an irreversible side effect of some pharmaceuticals which can contribute to tissue degeneration.
Objective To determine whether pharmaceutical glucocorticoids induce senescence in tenocytes.
Methods Features of senescence (β-galactosidase activity at pH 6 (SA-β-gal) and active mammalian/mechanistic target of rapamycin (mTOR) in cell cycle arrest) as well as the activity of the two main pathways leading to cell senescence were examined in glucocorticoid-treated primary human tenocytes. Evidence of senescence-inducing pathway induction in vivo was obtained using immunohistochemistry on tendon biopsy specimens taken before and 7 weeks after subacromial Depo-Medrone injection.
Results Dexamethasone treatment of tenocytes resulted in an increased percentage of SA-βgal-positive cells. Levels of phosphorylated p70S6K did not decrease with glucocorticoid treatment indicating mTOR remained active. Increased levels of acetylated p53 as well as increased RNA levels of its pro-senescence effector p21 were evident in dexamethasone-treated tenocytes. Levels of the p53 deacetylase sirtuin 1 were lower in dexamethasone-treated cells compared with controls. Knockdown of p53 or inhibition of p53 activity prevented dexamethasone-induced senescence. Activation of sirtuin 1 either by exogenous overexpression or by treatment with resveratrol or low glucose prevented dexamethasone-induced senescence. Immunohistochemical analysis of tendon biopsies taken before and after glucocorticoid injection revealed a significant increase in the percentage of p53-positive cells (p=0.03). The percentage of p21-positive cells also tended to be higher post-injection (p=0.06) suggesting glucocorticoids activate the p53/p21 senescence-inducing pathway in vivo as well as in vitro.
Conclusion As cell senescence is irreversible in vivo, glucocorticoid-induced senescence may result in long-term degenerative changes in tendon tissue.
- Corticosteroids
- Tendinitis
- Chondrocytes
- Fibroblasts
- Inflammation
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/
Statistics from Altmetric.com
Introduction
Musculoskeletal pain is one of the most common complaints of patients seeking general practitioner consultations. Chronic musculoskeletal pain arising from conditions such as arthritis and rotator cuff disease is often particularly difficult to treat. When oral analgesics prove ineffective, local injection with synthetic glucocorticoids is frequently used. In the UK, over 500 000 intra-articular glucocorticoid injections are administered per year in the primary care setting alone.1
Glucocorticoids are used to treat pathologies affecting a range of different tissues. Despite the prevalence of glucocorticoid use, there is concern glucocorticoids may have unwanted side effects.2–6 Glucocorticoids have been shown to induce cell cycle arrest and inhibit normal cell activity.7–10 However, it is not known whether these are transient effects quickly reversed once the glucocorticoid has dissipated from the tissue or whether the effects are more persistent.
Cell cycle arrest can lead to either quiescence or senescence. Quiescence is a reversible process leading to a transient state of cell dormancy. Senescence in contrast results in an irreversible change in normal cell activity. First observed in aged cells, cell senescence can also occur in young cells usually as a consequence of activation of either/both of the p38/p16INK4a or p53/p21cip stress-responsive signalling pathways. Senescent cells are hypersecretory, producing abnormal extracellular matrix as well as growth factors, cytokines and matrix-degrading enzymes.11 ,12 Many of these molecules are implicated in the pathogenesis of tendinopathy.13 ,14 Determining whether quiescence or senescence prevails following glucocorticoid treatment is therefore important for understanding the implications of glucocorticoid-induced cell cycle arrest.
Whether cell cycle arrest leads to quiescence or senescence hinges on the activity of mammalian/mechanistic target of rapamycin (mTOR).15 ,16 mTOR is the catalytic subunit of two enzyme complexes: mTORC1 and mTORC2. mTORC1 drives both cell proliferation and cell growth in part by phosphorylating the ribosomal protein S6 kinases.17–19 In quiescent cells, mTOR is inactive. However in senescent cells, mTOR remains active.15 As senescent cells are in cycle arrest, they are impervious to the pro-proliferative effects of mTOR but are responsive to its growth stimulatory effects. As a result, senescent cells continue to grow and appear enlarged compared with their non-senescent counterparts.20 Senescence-associated hypertrophy leads to increased lysosomal number and activity.21 It is this feature which is exploited in one of the most widely used assays to identify senescence—the activity of lysosomal β-galactosidase (SA-β-gal).22
The aims of the present study were to determine whether glucocorticoids induce quiescence or senescence in tenocytes in vitro, to identify the signalling pathways controlling this cell fate decision and to confirm whether the same cell phenotype results in tendon tissue following local injection with glucocorticoids.
Methods
Cell culture
Human tenocytes were obtained by explant culture of hamstring tendon used in repair of ruptured anterior cruciate ligament as previously described.10 Osteoblasts were obtained by explant culture of bone chips from osteoarthritic patients and chondrocytes by collagenase digestion of osteoarthritic knee cartilage. For more details, refer online supplementary information.
RNAi-mediated gene silencing
Gene knockdown was achieved using lipofectamine RNAimax (Life Technologies, Paisley, UK) following the manufacturer's instructions (for details refer online supplementary information). The transfection protocol did not alter cellular response to dexamethasone treatment (see online supplementary figure S1).
Adenoviral-mediated gene transduction
Tenocytes were infected with either a sirtuin 1 (adSirt1) or a green fluorescent protein (adGFP) adenoviral vector (refer online supplementary information for details) at MOI35 using Xtremegene HP (Roche Diagnostics Ltd, Burgess Hill, UK) following the manufacturer's instructions. The transduction protocol did not alter cellular response to dexamethasone treatment (see online supplementary figure S1).
Western blotting
Cells were sonicated in standard lysis buffer containing protease and phosphatase inhibitors. Western blots were carried out according to standard protocols10 and proteins visualised using Pierce WestDura detection reagents (ThermoScientific, Rockford, Illinois, USA) using a Chemi Doc-It Imaging System with Biochemi HR camera (UVP, Upland, California, USA).
Real-time reverse transcription polymerase chain reaction (RT-PCR)
cDNA was prepared using a cells-to-cDNA kit as per the manufacturer's instructions (Ambion, Austin, Texas, USA). Samples without reverse transcriptase and no-template controls served as negative controls. Real-time quantitative polymerase chain reaction (qPCR) reactions were performed using a Corbett Rotor-Gene 3000, a QuantiTect SYBR Green PCR kit and QuantiTect Primer Assays (Qiagen, Crawley, UK). All samples were run in duplicate with a coefficient of variation between duplicates of <1.0 cycle. Analysis was carried out using the delta-delta cT method.23
Senescence-associated β-galactosidase staining
SA-β-Gal activity was determined as previously described22 with minor modifications (see online supplementary information). Cells were visualised using a BX40 Olympus microscope with a DP70 camera (Olympus Life Science Europa Gmbh, Hamburg, Germany) with Safranin O as a counterstain. The number of β-galactosidase-positive cells and the total number of cells were counted in 10 fields of view (100×) per sample.
Pre-glucocorticoid and post-glucocorticoid injection tendon biopsies
Ethical approval for the study was granted by the local research ethics committee (Oxfordshire REC B, ref: 09/H0605/111). Supraspinatus tendon biopsies were taken from five patients undergoing subacromial glucocorticoid injection for rotator cuff tendinopathy (refer table 1 for patient details). All patients had subacromial impingement pain, ‘painful arc’ and positive Hawkins’ and Jobe's tests.24–26 Structural integrity of the rotator cuff was assessed by ultrasound. Exclusion criteria were full-thickness rotator cuff tear, previous significant shoulder trauma, systemic steroid use or tobacco smoking, previous or current shoulder problems other than rotator cuff tendinopathy, previous shoulder surgery, local glucocorticoid injections in the last 6 weeks or a significant systemic medical condition, for example, diabetes or a rheumatological condition. Bonar scores as described in ref. [27] were used to compare the degree of tendinopathy pre-glucocorticoid and post-glucocorticoid injection (table 1).
Pre-glucocorticoid and post-glucocorticoid injection biopsy study patient details
Longitudinal tendon biopsies were taken at a consistent location 5–10 mm posterior to the rotator interval under ultrasound guidance using a 14G spring-loaded core biopsy needle. Biopsies were taken immediately prior to and 7 weeks following ultrasound-guided injection of 40 mg Depo-Medrone into the subacromial bursa. Biopsies were immediately placed into 10% buffered formalin, subsequently wax embedded and sectioned (5 μm).
Immunohistochemistry
Immunohistochemistry was performed using an EnVision FLEX High-pH kit and an AutostainerLink48 as per the manufacturer's instructions (Dako, Ely, UK). p53 and p21 antibodies (refer online supplementary information) were used at dilutions of 1:50 and 1:100, respectively.
Statistical analysis
All in vitro experiments were repeated at least three times using different tissue donors for each experimental replicate. The results for in vitro experiments were analysed by one-way analysis of variance (ANOVA) with post hoc Tukey test or by t test if only two conditions were being tested. Data obtained from immunohistochemistry on patient specimens were analysed using the non-parametric Wilcoxon matched pairs t test. All data were analysed using Prism V.5.0b (GraphPad Software, La Jolla, California, USA). p Value ≤0.05 was considered statistically significant. The results are expressed as mean±SD.
Results
Glucocorticoid treatment results in cell morphological changes consistent with a senescent phenotype
To treat tendinopathy, typically 40 mg of glucocorticoid suspended in 1–10 ml of saline is injected into the bursa (equating to 10–100 mM). The glucocorticoid concentration to which tenocytes are exposed following local injection is difficult to assess and will depend on the perfusibility of the tendon tissue and the proximity of the tenocyte to the injection site. Following systemic administration, circulating glucocorticoid concentrations of approximately 1 μM have been observed.28
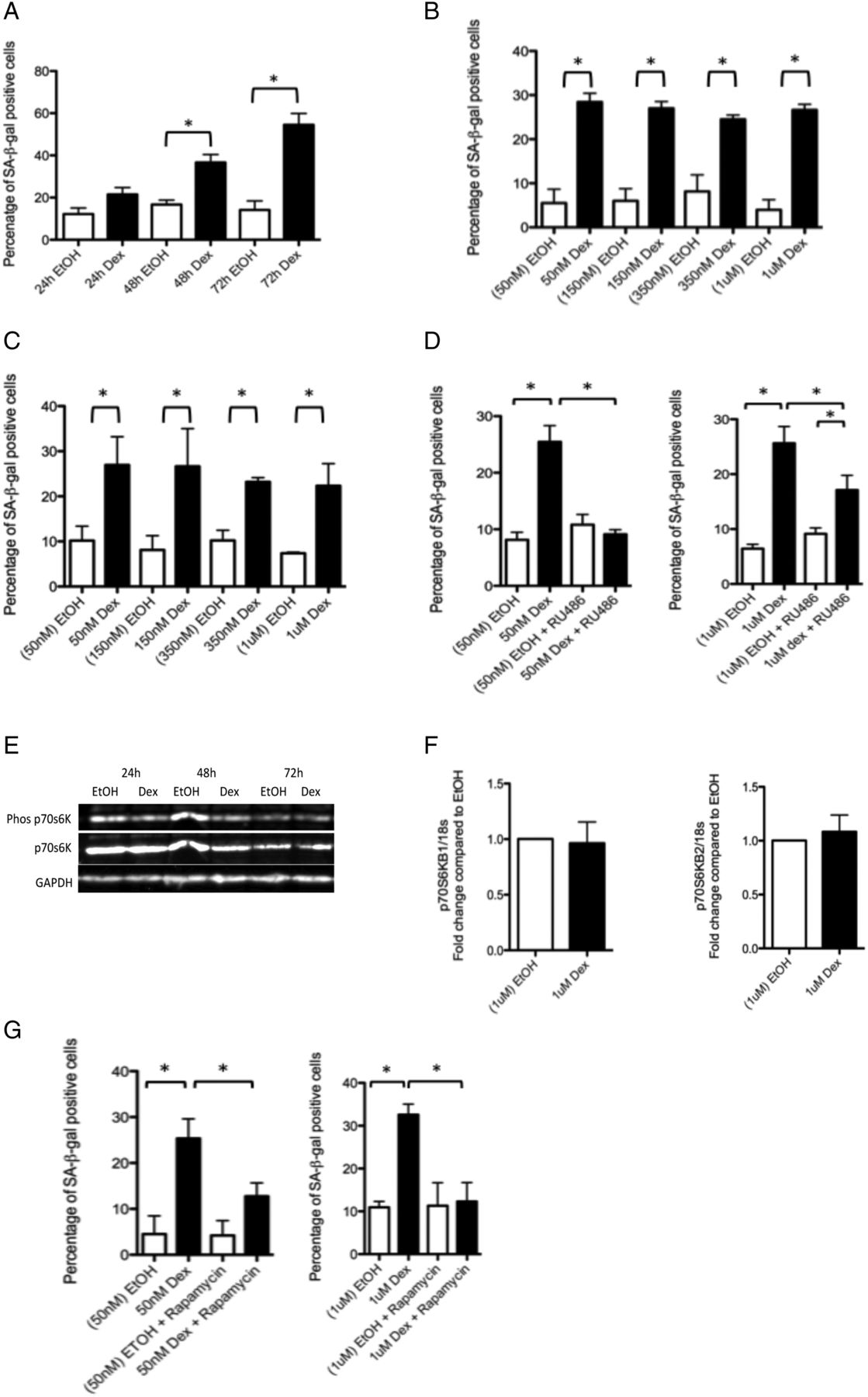
As an initial screen, we treated tenocytes with 1 μM dexamethasone. By 48 and 72 h post-treatment, a significant increase in the percentage of SA-β-gal-positive cells was observed (figure 1A, see online supplementary figure S2).

Glucocorticoids cause morphological and functional changes in tenocytes consistent with a senescent phenotype. (A) The percentage of SA-βgal-positive cells was significantly higher 48 and 72 h post-treatment of primary human tenocytes with 1 μM dexamethasone compared with tenocytes treated with ethanol (carrier) alone. There was no evidence of a dose effect of dexamethasone treatment on the percentage of SA-βgal-positive cells following (B) 3 days or (C) 7 days of treatment (n=3) compared with cells treated with the corresponding amount of ethanol carrier. (D) The percentage of SA-βgal-positive cells was significantly lower in cells treated for 72 h with dexamethasone (50 nM or 1 μM) and the glucocorticoid receptor (GR) antagonist RU486 (80 nM) compared with cells treated with dexamethasone alone, n=3. (E) Western blot showing levels of phosphorylated and total p70 S6 kinase, a substrate of the mTORC1 complex and an indicator of mTORC1 activity 24, 48 and 72 h post-dexamethasone (dex) treatment (1 μM) in primary human tenocytes. Western blots were conducted on protein lysates from cells obtained from three different tissue donors. Blot shown is representative of all experiments. (F) No difference in RNA levels of either of the two p70 S6 kinase subunits (p70S6KB1 and p70S6KB2) was evident in tenocytes treated with dexamethasone (1 μM) compared with ethanol-treated controls. (G) A significantly lower percentage of SA-βgal-positive cells was evident in cells treated for 72 h with dexamethasone (50 nM or 1 μM) and the mammalian/mechanistic target of rapamycin (mTOR) inhibitor rapamycin (rapa) compared with cells treated with dexamethasone alone, n=3. Statistically significant differences between ethanol (EtOH) and dexamethasone (Dex) and between dexamethasone with and without either the GR inhibitor (RU486) or mTOR inhibitor (rapamycin) are indicated by * (p<0.05).
In order to determine the effect of lower glucocorticoid doses, we treated tenocytes with 50nM, 150nM, 350 nM or 1 μM dex for 3 days. We found treatment with any of the concentrations tested resulted in an increased percentage of SA-β-gal-positive cells (figure 1B). Prolonged treatment for 7 days did not result in a further increase in the percentage of SA-β-gal-positive cells (figure 1C). Co-treatment with the glucocorticoid receptor (GR) antagonist RU486 (80 nM) for 72 h prevented the dex-induced increase in the percentage of SA-β-gal-positive cells, albeit more effectively, following 50 nM rather than 1 μM dexamethasone treatment (figure 1D).
mTOR remains active in glucocorticoid-treated tenocytes
We found no difference in levels of phosphorylated p70 S6K (figure 1E) or in RNA levels of either of the p70S6K subunits (figure 1F) between dex-treated tenocytes and controls. To confirm that mTOR was active following dex treatment, we co-treated cells with dex and the mTOR inhibitor rapamycin and found this led to a significant reduction in SA-β-gal-positive cells (figure 1G).
Glucocorticoids activate the p53 pathway
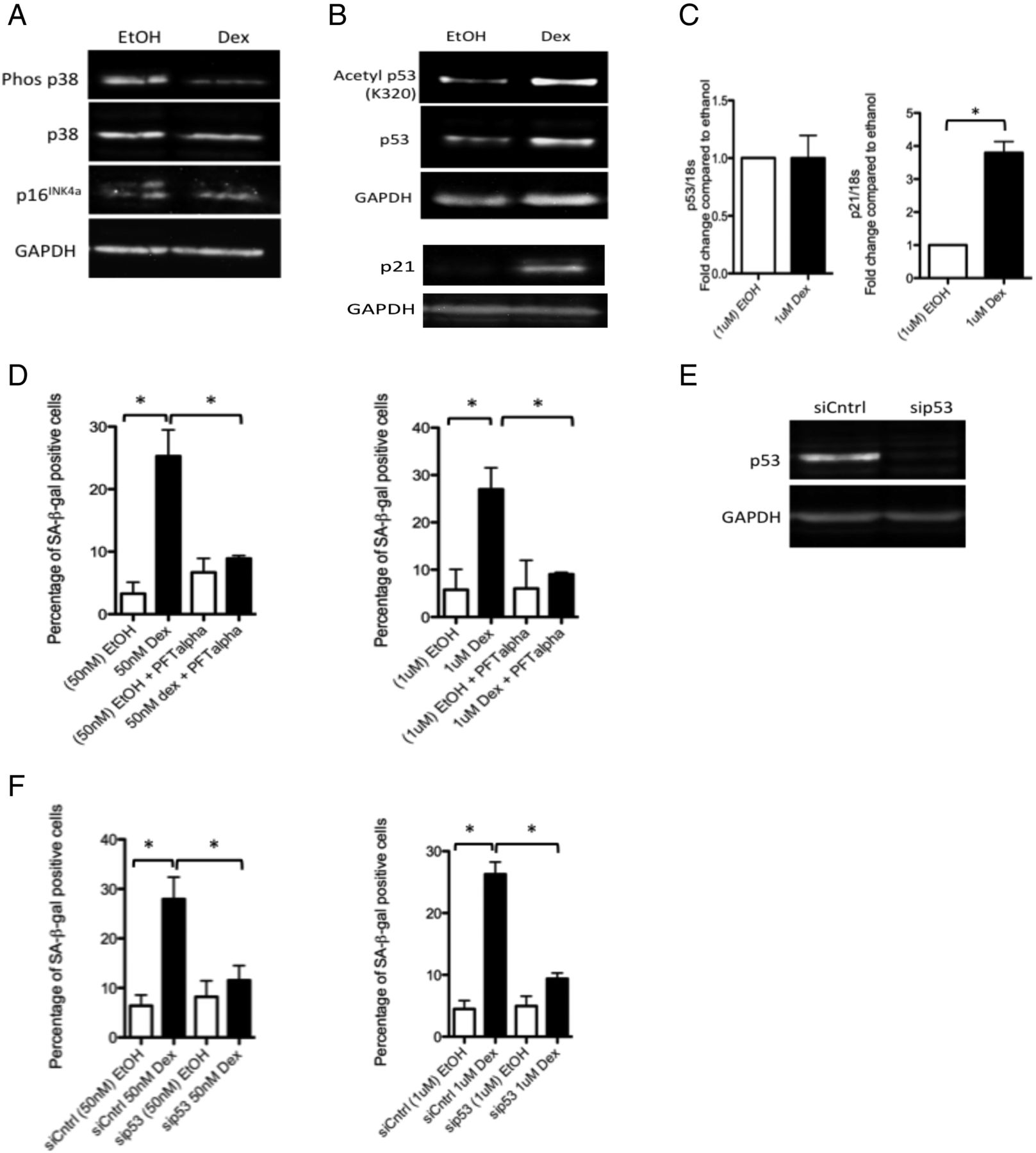
There are two major pathways leading to induction of cellular senescence: the p38 mitogen activated protein kinase (MAPK)/p16INK4a pathway and the p53/p21cip pathway.29 We found no evidence of an increase in levels of phosphorylated p38 MAPK or p16INK4a in dexamethasone-treated tenocytes compared with controls (figure 2A). In contrast, levels of acetylated p53 (figure 2B) (but not phosphorylated p53) (see online supplementary figure S3) were higher in glucocorticoid-treated tenocytes compared with controls. While there were no significant differences in p53 RNA levels, RNA levels of p21cip/waf1, a pro-senescence modulator of p53 activity, were markedly elevated post-glucocorticoid treatment (figure 2C).
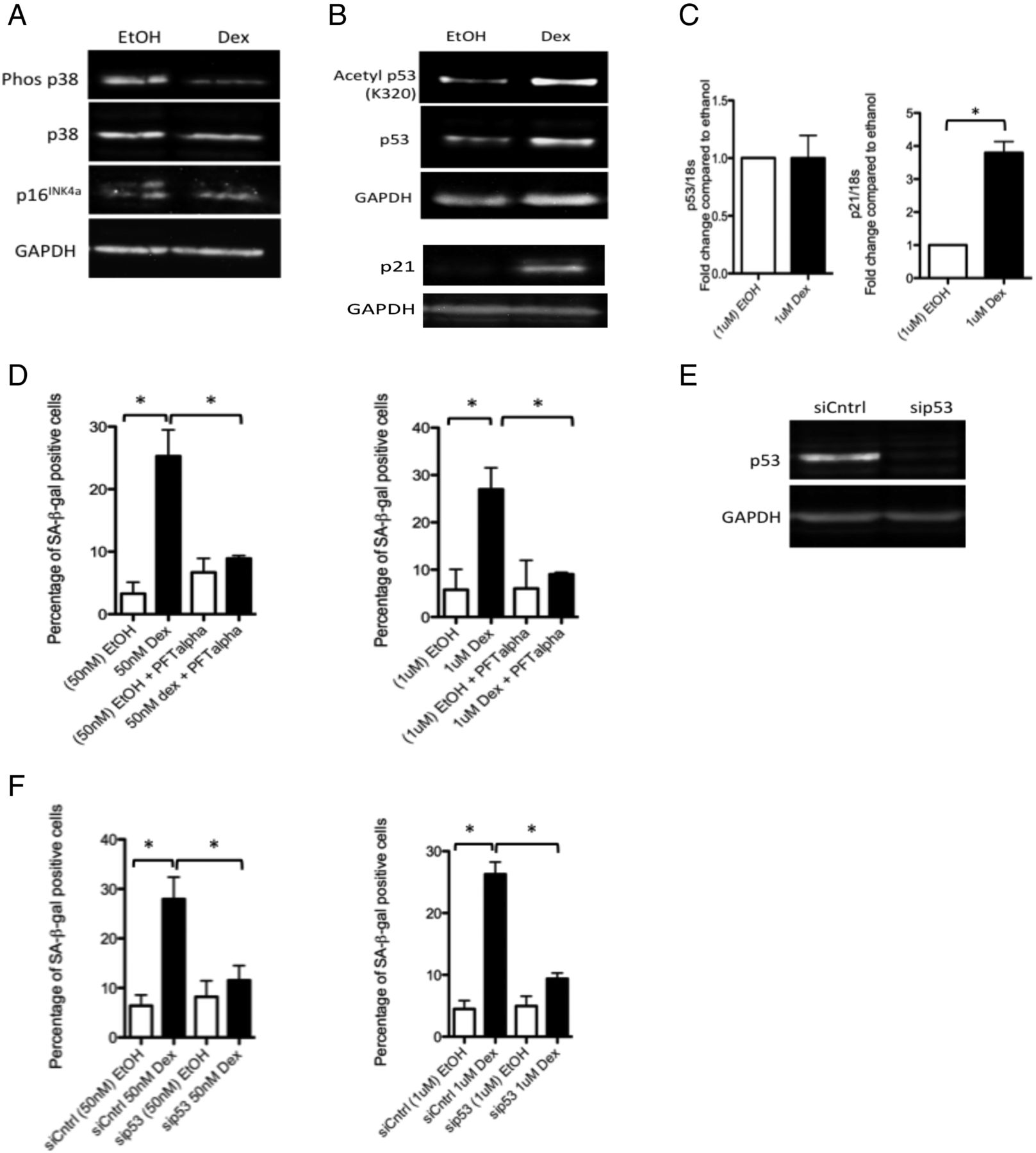
Glucocorticoids activate p53 signalling in tenocytes. Western blots showing levels of (A) phosphorylated and total p38 MAPK and p16INK4a and (B) acetylated (K320) and total p53 and p21cip/waf1 in lysates of primary human tenocytes following treatment with 1 μM dexamethasone or carrier (ethanol) for 48 h. Western blots were conducted on protein lysates from cells obtained from three different tissue donors. Blots shown are representative of all experiments. (C) No significant differences in RNA levels of p53 were apparent in tenocytes treated with 1 μM dexamethasone compared with ethanol-treated controls. However, RNA levels of p21 were significantly greater in tenocytes treated with dexamethasone compared with ethanol-treated controls. (D) The percentage of SA-βgal-positive cells was significantly lower in cells treated with dexamethasone (50 nM or 1 μM) and the p53 inhibitor pifithrin-α (PFT-α, 100 nM) compared with cells treated with dexamethasone alone, n=3. (E) Western blot demonstrating the effectiveness of p53 knockdown obtained using RNAi in primary human tenocytes. (F) The percentage of SA-βgal-positive cells was significantly lower in tenocytes in which p53 expression was knocked down (sip53) compared with dexamethasone-treated controls (siCntrl) (n=3). Statistically significant differences between ethanol (EtOH) and dexamethasone (Dex) and between dexamethasone with and without the p53 inhibitor (PFT-α) or with and without p53 siRNA (sip53) are indicated by * (p<0.05).
To confirm the involvement of p53 activity in glucocorticoid-induced senescence, we co-treated tenocytes with dexamethasone and the p53 inhibitor pifithrin-α (PFT-α, 100 nM) for 72 h. We found the number of SA-β-gal-positive cells was significantly lower in dexamethasone+inhibitor-treated cells compared with those treated with dexamethasone alone (figure 2D). Similarly, we found the number of SA-β-gal-positive cells was significantly lower in dexamethasone-treated cells in which p53 expression was knocked down using RNAi compared with dexamethasone-treated cells transfected with a non-targeting siRNA control (figure 2E, F).
Glucocorticoids inhibit expression of sirtuin 1
Reduced levels of the p53 deacetylase sirtuin 1 (sirt1) are often associated with p53-mediated senescence.30 We found a small but significant reduction in RNA levels of sirt1, but a marked reduction in protein levels in glucocorticoid-treated cells compared with carrier-treated controls (figure 3A,B). To determine whether sirt1 is involved in glucocorticoid-induced senescence, we overexpressed sirt1 in tenocytes using adenoviral-mediated gene delivery. We found the percentage of SA-β-gal-positive cells was significantly lower in dexamethasone-treated tenocytes infected with a sirt1-bearing adenoviral vector (adSirt1) compared with dexamethasone-treated tenocytes infected with a GFP-bearing control adenoviral vector (adGFP) (figure 3C,D).

Glucocorticoids inhibit sirtuin 1 expression in tenocytes. Both (A) RNA (as measured by real-time qRT-PCR 7 h post-treatment, n=3) and (B) protein (as determined by western blotting) levels of sirtuin 1 were reduced in dexamethasone-treated cells compared with carrier (ethanol)-treated controls. Western blots were conducted on protein lysates from cells obtained from three different tissue donors. Blot shown is representative of all experiments. (C) The percentage of SA-βgal-positive cells was significantly lower in dexamethasone-treated tenocytes infected with an adenoviral vector containing a sirtuin 1 overexpression construct (adSirt1) compared with dexamethasone-treated tenocytes infected with a control adenoviral vector expressing GFP (adGFP). (D) Western blot demonstrating the effectiveness of the adSirt1 virus to prevent dexamethasone-induced inhibition of sirtuin 1 expression. Statistically significant differences between ethanol (EtOH) and dexamethasone (Dex) and between dexamethasone with and without sirt1 overexpression (adSirt1) are indicated by * (p<0.05).
Prevention of the glucocorticoid-mediated inhibition of sirtuin 1 expression protects against senescence induction
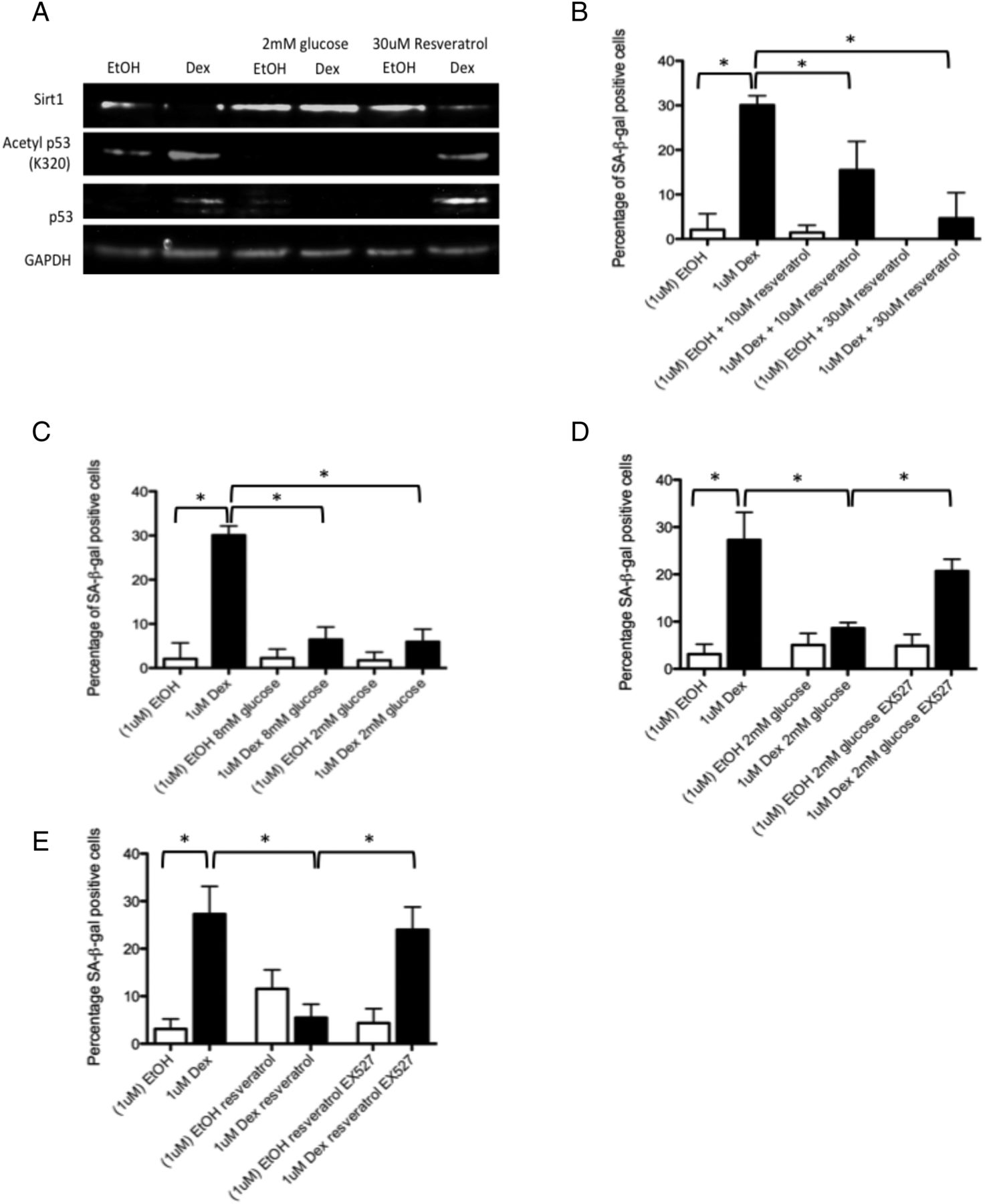
sirt1 is activated by caloric restriction and by the plant-derived phenol resveratrol. To simulate caloric restriction in vitro, we cultured cells in low (2 mM) glucose media. We found protein levels of sirt1 were higher, and levels of acetylated p53 lower, in tenocytes co-treated with dexamethasone and resveratrol or in dexamethasone-treated tenocytes grown in low glucose media compared with cells treated with dexamethasone alone and grown in standard (17.5 mM glucose) culture media (figure 4A). The number of SA-β-gal-positive cells was also significantly lower in cells co-treated with dexamethasone and resveratrol or treated with dexamethasone in low glucose media compared with cells treated with dexamethasone alone indicating these treatments can protect against glucocorticoid-induced senescence in vitro (figure 4B, C). In order to confirm the involvement of sirt1 activity in the protective effect of these two treatments against glucocorticoid-induced senescence, we used a specific sirt1 inhibitor (100 nM EX527). The percentage of SA-β-gal-positive cells was significantly higher in dexamethasone-treated cells co-treated with the sirt1 inhibitor and cultured in low glucose (2 mM) media (figure 4D) or treated with resveratrol (30 μM) (figure 4E) compared with cells grown under the same conditions without the sirt1 inhibitor.

Protective effect of low glucose, low serum and resveratrol on glucocorticoid-induced senescence (A) Protein levels of sirtuin 1 were higher in tenocytes treated with dexamethasone (1 μM) and cultured in low glucose (2 mM) media or co-treated with resveratrol (30 μM) compared with tenocytes treated with dexamethasone and cultured in standard growth media (17.5 mM glucose). Levels of acetylated p53 (lys320) were lower in dexamethasone-treated cells cultured in low glucose (2 mM) media or co-treated with resveratrol compared with tenocytes treated with dexamethasone and cultured in standard growth media (17.5 mM glucose). Western blots were conducted on protein lysates from cells obtained from three different tissue donors. Blots shown are representative of all three experiments. The percentage of SA-βgal-positive cells was significantly lower in dexamethasone-treated cells (B) co-treated with resveratrol (10 or 30 μM) or (C) cultured in low glucose (2 or 8 mM) media; however, no significant difference in the percentage of SA-βgal-positive cells was observed following addition of EX527, a sirtuin 1 inhibitor, in dexamethasone-treated cells (1 μM) cultured in (D) 2 mM glucose or (E) co-treated with resveratrol (30 μM) compared with cells treated with dexamethasone alone and cultured in standard (17.5 mM) growth media (n=3). Statistically significant differences between ethanol (EtOH) and dexamethasone (Dex) and between dexamethasone with and without either resveratrol or 2 mM glucose are indicated by * (p<0.05).
Evidence of p53 pathway activation following local injection of glucocorticoids in patients with rotator cuff disease
Tendon biopsies were taken from five patients with rotator cuff disease immediately preceding and 7 weeks following glucocorticoid injection. Using immunohistochemistry, we found the percentage of p53-positive cells was significantly higher (p=0.03) post-injection compared with pre-injection (figure 5A). The percentage of p21-positive cells also tended to be higher post-injection (p=0.06) (figure 5B). There was no significant difference in the degree of tendinopathy as assessed by Bonar score between pre-injection and post-injection tissue biopsies (see online supplementary figure S4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The p53/p21 pathway is activated in vivo following local injection with glucocorticoids. Tendon biopsy samples were taken from five patients with rotator cuff tendinopathy immediately prior to subacromial injection with the synthetic glucocorticoid, Depo-Medrone, and compared with subsequent biopsy specimens taken from the same patients 7 weeks following glucocorticoid injection. Using immunohistochemistry, (A) the percentage of p53-positive cells was significantly greater in tissue samples post-injection compared with pre-injection. (B) A non-statistically significant increase in the percentage of p21-positive cells was observed in the post-injection tissue samples compared with pre-injection tissue samples. Representative photomicrographs are shown for tissue samples from two patients. p53-positive cells or p21-positive cells were detected using horseradish peroxidase-conjugated secondary antibodies and diaminobenzadine. Positive cells appear brown. Tissue was counterstained with haematoxylin (blue) to allow quantification of total cell numbers. Statistically significant differences are indicated by * (p<0.05).
Glucocorticoids also induce senescent-like changes in primary human chondrocytes and osteoblasts
To determine whether other cell types also undergo senescence-like changes following dexamethasone treatment, we treated primary human chondrocytes and primary human osteoblasts with dexamethasone (1 μM) for 72 h. We found the percentage of SA-β-gal-positive cells was significantly higher in both cell types following dexamethasone treatment (see online supplementary figure S5).
Discussion
In the present study, we have demonstrated that glucocorticoids induce senescence in tenocytes in vitro by the p53/p21 pathway. We have also shown that the p53/p21 pathway is upregulated in tendon tissue 7 weeks after local injection of glucocorticoids suggesting that glucocorticoids also induce senescence in vivo. These findings represent a previously undescribed mechanism of action of glucocorticoids likely to have important clinical implications.
We, and others, have previously reported that glucocorticoid treatment of tenocytes leads to inhibition of cell proliferation7 ,9 ,10 but whether this results in cell quiescence or senescence was unknown. One of the critical features distinguishing a senescent cell from a quiescent cell is that mTOR remains active in the former. We found no change in levels of phosphorylated p70 S6kinase, a substrate of the mTORC1 enzyme, following glucocorticoid treatment of tenocytes in situ suggesting mTOR remained active in glucocorticoid-treated cells. Consistent with this finding, SA-β-gal activity was greater in glucocorticoid-treated tenocytes compared with controls. Inhibition of mTOR with rapamycin markedly reduced the number of SA-β-gal-positive cells following glucocorticoid treatment. Taken together, these results confirm that glucocorticoids induce senescence in primary human tenocytes.
Circulating endogenous glucocorticoids are mostly protein bound and as such are inactive. Local injection of glucocorticoids results in a large influx of unbound and therefore active glucocorticoid into the joint. Similarly, almost all of the glucocorticoids administered in our in vitro experiments would have been ‘free’, that is, available for receptor binding. In the present study, we saw no evidence of a dose effect of glucocorticoids on senescence induction. It is likely that the large excess of unbound glucocorticoid provided even by our lowest glucocorticoid dose fully occupied receptor sites, resulting in maximal receptor activation.
There are two major signalling pathways which can lead to induction of cellular senescence: the p38 MAPK/p16INK4a pathway and the p53/p21cip pathway (reviewed in ref. 29). We found no evidence of activation of the p38 MAPK/p16INK4a pathway in glucocorticoid-treated tenocytes. In contrast, we found levels of acetylated p53 were higher in glucocorticoid-treated tenocytes compared with carrier-treated controls indicating glucocorticoids activate p53 signalling in tenocytes. Consistent with our results, Crochemore et al31 found that GR activation enhanced p53 activity in neural cells. Acetylation of p53 on K320 promotes the transcription of pro-survival (as opposed to pro-apoptosis) p53-regulated genes. It also stabilises the p53 protein by preventing its nuclear export.32 This stabilisation of p53 may be responsible for the increase in total p53 protein levels also observed in glucocorticoid-treated cells in the present study.
p53 activation can lead to either promotion of apoptosis or induction of senescence. p21cip is a cell cycle controller which is critical for determining the outcome of p53 activation. p21cip induces cell cycle arrest and inhibits the pro-apoptotic activity of p53 channelling p53 activity towards senescence induction.33 Glucocorticoids have previously been shown to directly stimulate p21cip transcription.34 In the present study, we observed increased p21cip expression following glucocorticoid treatment of tenocytes indicating the pro-senescence functionality of p53 was activated by glucocorticoid treatment. Confirming the involvement of p53 signalling in glucocorticoid-induced senescence, we found inhibiting p53 activity or knockdown of p53 expression in glucocorticoid-treated tenocytes resulted in significantly fewer SA-β-gal-positive cells.
p53 is deacetylated by sirt1.35 ,36 Reduced sirt1 levels and/or activity are frequently associated with increased p53 acetylation and increased p53-mediated senescence.37 ,38 We found glucocorticoid treatment resulted in a significant reduction in both RNA and protein levels of sirt1 in vitro. To our knowledge, this is the first report describing glucocorticoid-mediated regulation of sirt1 expression. Caloric restriction and the plant-derived phenol resveratrol increase both the expression39 and the activity of sirt1.40–42 In the present study, we found lowering the glucose content of the cell culture medium (to emulate caloric restriction) or treatment with resveratrol protected against the glucocorticoid-induced reduction in sirt1 levels, partially ameliorated the increase in acetylated p53 levels and led to significantly fewer SA-β-gal-positive cells. The inhibitory effect of low glucose and resveratrol on glucocorticoid-induced senescence was abolished by the addition of a sirt1 inhibitor demonstrating that the activation of sirt1 was largely responsible for their protective effect. In concurrence with these results, we found overexpression of sirt1 in tenocytes using adenoviral-mediated gene transduction also protected against glucocorticoid-induced senescence.
To determine whether glucocorticoid treatment also induces senescence in patients, we compared supraspinatus biopsies taken before and after subacromial glucocorticoid injection in five patients with rotator cuff tendinopathy. Using this set of biopsy specimens, we have previously found that the percentage of cells positive for the proliferation marker mib-1 was significantly lower, but the percentage of apoptotic cells was not significantly different in post-injection biopsies compared with pre-injection biopsies (manuscript in preparation). In the present study, we found the percentage of p53-positive cells was significantly higher in post-injection biopsies compared with pre-injection biopsies (p=0.03). The percentage of p21-positive cells also tended to be higher post-injection compared with pre-injection (p=0.06). These data provide compelling evidence that the p53/p21 pathway is upregulated following glucocorticoid injection in patients suggesting that glucocorticoids also induce senescence in vivo.
Synthetic glucocorticoids are extensively used to treat painful and inflammatory conditions in a variety of tissues. In vitro, we found the percentage of SA-β-gal-positive cells was also higher following glucocorticoid treatment of primary human osteoblasts and chondrocytes suggesting glucocorticoids may induce senescence in a range of different cell types.
The loss of normal functionality associated with senescence has been implicated in age-related disease and degeneration.43 ,44 Given the apparent irreversible nature of senescence, glucocorticoid-induced senescence is likely to have long-term detrimental consequences for tissue. That we observed a marked increase in p53 expression 7 weeks following glucocorticoid injection supports the notion that the effects of local glucocorticoid injection on tendon tissue are not transient. Glucocorticoid-induced senescence may exacerbate the underlying tissue pathology responsible for the pain for which the glucocorticoids are prescribed to treat.
Acknowledgments
The authors gratefully acknowledge Mr Chethan Jayadev, Mr Ben Davies, Professor Andrew Price and the Oxford Musculoskeletal Biobank (OMB), University of Oxford, UK for collection of human hamstring, cartilage and bone tissue. Sincere thanks to Dr Benjamin Dean and Dr Sarah Franklin for tissue sectioning and help with immunohistochemistry. To research nurses, Mrs Kim Wheway and Mrs Bridget Watkins for help with collection of biopsies pre-glucocorticoid and post-glucocorticoid injection. Special thanks also to the patients who donated their time and tissue to make this study possible.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
- Data supplement 2 - Online figures
Footnotes
-
Handling editor Tore K Kvien
-
Contributors RP was involved in conception and design of the study, acquisition, analysis and interpretation of data and writing of the manuscript. AW was involved in data acquisition and writing of the manuscript. RM was involved in conception and design of the biopsy study, collection of biopsy specimens and writing of the manuscript. SS was involved in data acquisition and revision of the manuscript. AC was involved in conception and design of the biopsy study, biopsy collection and critical review of the manuscript. PH was involved in study design and review of the manuscript.
-
Funding This study was funded by Arthritis Research UK (grant no. 19482) and the NIHR Oxford Musculoskeletal Biomedical Research Unit. The extent of involvement of both sponsors in this study was purely financial.
-
Competing interests None.
-
Ethics approval Oxfordshire REC B.
-
Provenance and peer review Not commissioned; externally peer reviewed.