Article Text

Abstract
Objective Notch signalling pathways are critical for angiogenesis and endothelial cell (EC) fate; however the mechanisms regulating these processes in the inflamed joint remain to be elucidated. Here, we examine whether Notch signalling mediates vascular endothelial growth factor (VEGF) and angiopoietin 2 (Ang2)-induced vascular function.
Methods Notch-1 intracellular domain (Notch-1 IC), Notch-4 IC, Delta-like-ligand 4, Hes-related transcriptional repressors-1 and 2 (Hrt-1, Hrt-2) mRNA and/or protein expression was measured by Real-time PCR and/or western blot. VEGF/Ang2 induced EC function was assessed using transwell invasion chambers, matrigel tube formation assays and wound repair scratch assays ± Notch-1 siRNA or an γ-secretase inhibitor N-(N-(3,5-Difluorophenacetyl-L-alanly))-S-phenylglycine-t-Butyl Ester (DAPT) in RA synovial explants or human microvascular EC. Interleukin (IL)-6 and IL-8 were measured by ELISA and MMP2 and 9 by gelatine zymography.
Results Notch-1 IC and Notch-4 IC protein expressions were demonstrated in RA and psoriatic arthritis synovial biopsies, with minimal expression observed in Osteoarthritis (OA). VEGF and Ang2 induced Notch-1 IC/ Notch-4 IC protein expression in synovial explant cultures and human microvascular EC levels were further potentiated by VEGF/Ang2 stimulation in combination. Notch-1, Delta-like-ligand 4, and Hrt-2 mRNA expression were significantly induced by VEGF and Ang2 alone and in combination. Furthermore VEGF/Ang2-induced EC invasion, angiogenesis and migration were inhibited by Notch-1 siRNA or DAPT. Conditioned media from VEGF/Ang2 stimulated RA synovial explants induced EC tube formation, an effect that was inhibited by DAPT. Finally, DAPT significantly decreased VEGF/Ang2 induced IL-6, IL-8, MMP2 and 9 expressions in RA synovial explants.
Conclusions Notch-1 mediates VEGF/Ang2-induced angiogenesis and EC invasion in inflammatory arthritis.
- Inflammation
- Rheumatoid Arthritis
- Autoimmune Diseases
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/3.0/ and http://creativecommons.org/licenses/by-nc/3.0/legalcode
Statistics from Altmetric.com
Introduction
Inflammatory arthritis (IA) is a progressive autoimmune disease affecting 1% of the population.1 Dysregulated angiogenesis is an early event in IA facilitating synovial membrane (SM) vessel invasion, persistent leukocyte infiltration and lining layer hyperplasia, capable of destroying adjacent cartilage and bone.2–4 Vascular endothelial growth factor (VEGF), Angiopoietin (Ang), platelet derived growth factor and transforming growth factor-β all regulate vessel stability and induce fibroblast invasion.4–10 Furthermore, studies demonstrate similar dysfunctional microvessels in the papillary dermis of skin in Psoriatic arthritis (PsA) patients associated with differential expression of VEGF, Ang1/2, MMP-9 and E-selectin.11–14
VEGF stimulates endothelial cell (EC) proliferation, permeability, migration and promotes EC survival.3 ,15 Ang, a family of EC specific factors are also fundamental in vessel formation.16–21 Complementary action between VEGF and Ang controls vessel stability and maturation.19–21 Ang1 knockout mice are embryo lethal, due to absent capillary sprouts and vessel wall deformity.16 Ang1 overexpression increases EC/pericyte interactions resulting in stable vasculature while angiopoietin 2 (Ang2) antagonises Ang1 on invading vascular sprouts, blocking stabilisation.18–20 Differential expression of Ang1, Ang2 and VEGF has been demonstrated in synovial tissue/cells and are associated with differential pathogenic outcomes.5 ,8 ,22 ,23 Expression of Ang1, Ang2 and their receptor Tie2 are significantly increased in progression of collagen induced arthritis (CIA), and Tie2 blockade ameliorates bone destruction.24 ,25 Recently, we demonstrated that Tie2 mediates Toll-like receptor-2 induced angiogenesis in Rheumatoid arthritis (RA).26
The Notch signalling pathway plays a pivotal role in vascular development, cell-cell communication, cell fate decisions.27–30 Notch is also critical for EC-pericyte interactions and vascular network remodelling.28–33 Four Notch receptors are described in mammals with ligands encoded by Jagged-1, 2 and Delta-like 1, 3, 4 (DLL-1, DLL-3, DLL-4) genes.34–41 Cleavage of Notch receptors releases Notch intracellular domain which translocates into the nucleus,39 ,40 regulating downstream target genes, Hrt (Hes-related transcriptional repressors) and Hes (Hairy/Enhancer of Split).28 ,41 Previous studies have shown Notch and/or its receptors in inflamed SM and synoviocytes.42 ,43 Jagged-1 modulates CIA by regulating T cell responses.44 Notch-1 can mediate TNFα-induced synoviocyte proliferation,42 ,45 and Notch-3 and DLL-1 mediate collagen-specific T-cell activation and altered T helper cells responses.46 However, the mechanisms by which Notch signalling regulates angiogenesis in the inflamed joint remain to be elucidated. In this study we examine if Notch signalling mediates VEGF/Ang2 induced angiogenesis in the inflamed joint using ex vivo synovial explant cultures and microvascular ECs.
Material and methods
Patient recruitment and arthroscopy
Twenty-nine subjects (10M: 19F) were recruited to this study (RA=10; PsA=10; OA=9). Synovial tissue biopsies were obtained at arthroscopy as previously described.47 Patients with RA and PsA, fulfilled the American College of Rheumatology48 and Classification Criteria for Psoriatic Arthritis (CASPAR)49 criteria. The median age of the RA patients was 52.84 (27.26–80.22) years, the PsA 64.39 (32.27–80.58) years and OA 55.89 (37.22–77.21) years. The median DAS28 for RA patients was 4.565 (1.75–6.23), for PsA 3.625 (2.1–4.92) and OA 4.025 (3.25–4.05). Fifty per cent of inflammatory patients (RA/PsA) were naive for disease modifying antirheumatic drugs and steroids, others had failed at least one disease modifying antirheumatic drugs. Following institutional approval by the St. Vincent's University Hospital medical research and ethics committee, all patients gave written informed consent. All treatment was fully compliant with the Helsinki Declaration.
Ex vivo synovial explant culture
To examine the effect of VEGF/Ang2 alone and in combination on Notch-1 intracellular domain (Notch-1 IC) and Notch-4 IC expression an ex vivo whole synovial tissue explant model was established.26 RA/PsA synovial explant tissue was sectioned into 96-well-plates (Falcon, Franklin Lakes, New Jersey, USA) in Roswell Park Memorial Institute (RPMI) 1640 supplemented with streptomycin (100 units/ml) and penicillin (100 units/ml) and cultured with VEGF (20 ng/ml)50 (R&D systems, Abingdon, UK), Ang2 (250 ng/ml)51 (R&D systems) alone and in combination for 24 h at 37°C in 5% CO2. Supernatants were harvested and tissue was snap frozen for protein analysis.
Culture of HMVEC
Human microvascular ECs (HMVEC) (Lonza, Waterville, Inc, California, USA), were grown in endothelial basal medium (EBM) supplemented with 5% fetal calf serum (FCS), 0.5 ml human epidermal growth factor (hEGF), 0.5 ml hydrocortisone, 0.5 ml gentamicin, 0.5 ml bovine brain extract (Lonza) and used for experiments between passages 3–8. For Notch-1 IC and Notch-4 IC protein expression, HMVEC were grown to confluence, then cultured in serum reduced EBM for 24 h (1% FCS) before stimulation with VEGF (20 ng/ml) alone and in combination with Ang2 (250 ng/ml) for a further 24 h.
Western blot analysis
Synovial tissue and HMVEC protein lysates were prepared as previously described.52 Proteins from synovial tissue lysates and HMVEC were resolved on sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) prior to transfer onto nitrocellulose membrane (Amersham Biosciences, Buckinghamshire, UK). Rabbit polyclonal anti Notch-1 or Notch-4 (Millipore, Temecula, California, USA) were used as previously described.52 Blots were developed using enhanced chemiluminescence (ECL) (Pierce, Rockford, Illinois, USA) and signal intensity was quantified by densitometry using the Electrophoresis Documentation and Analysis System (EDAS) 120 system (Kodak, Rochester, New York, USA). Full-length Notch-1 (300 kDa) and extracellular fragment (200 kDa) were also observed in addition to cytoplasmic domain (120 kDa) as per the manufacturer's instructions.
Immunohistochemistry
Immunohistochemical analysis for Notch-1 and Notch-4 in RA, PsA and OA tissue was performed as previously described.52 Briefly sections were incubated with primary antibodies against rabbit-polyclonal Notch-1, Notch-4 (Millipore) and isotype matched rabbit-polyclonal IgG control (DAKO, UK) at room temperature for 1 h. Colour was developed in solution containing diaminobenzadine-tetrahydrochloride (Sigma), 0.5% H2O2 in phosphate buffered saline (PBS) buffer (pH 7.6). Slides were counterstained with haematoxylin and mounted. Slides were analysed using a well-established semiquantitative scoring method.52 ,53
Notch-1 siRNA gene silencing studies
For each 25 cm2 flask of HMVEC transfected, 5 µl of 20 pmol gene-specific siRNA duplexes (Notch-1 or Scramble) and 5 µl of Lipofectamine 2000 Reagent (Invitrogen, BioSciences Ltd., Ireland) were mixed gently with 0.99 ml serum/antibiotic-free Optimal Minimal Essential Medium (OPTI-MEM) (Invitrogen) and incubated at room temperature for 20–30 min in the dark. The combination was mixed with full (5% FCS) EBM, added to cells and incubated overnight. The siRNA duplexes for Notch-1 (5459NM-017617) and Scrambled control (a nonsense siRNA of the target sequence) were from Sigma.
γ-Secretase inhibitor (DAPT) studies
For γ-secretase inhibitor study, biopsies were sectioned into 96-well-plates and HMVEC were plated in 48-well-plates. Cells were cultured with VEGF (20 ng/ml), Ang2 (250 ng/ml) alone and in combination in the presence or absence of N-(N-(3,5-Difluorophenacetyl-L-alanly))-S-phenylglycine-t-Butyl Ester (DAPT) (Sigma) used at between 1–50 µM, or Dimethyl sulfoxide (DMSO) vehicle control. The supernatants (conditioned media) were harvested for cytokines and tube formation analysis.
mRNA extraction and analysis
HMVEC were grown to confluence, cultured in serum reduced EBM for 24 h (1% FCS) and then stimulated with VEGF (20 ng/ml) alone and in combination with Ang2 (250 ng/ml) for 3, 6 and 24 h. Total RNA was isolated using RNeasy Mini Kit (Qiagen, Crawley, UK) according to the manufacturer's specifications. Purity of RNA was measured and samples with a ratio over 1.8 (260 : 280 nm) used in subsequent experiments. Total RNA (1–2 µg) was reverse transcribed to cDNA. Relative quantification of gene expression was analysed with preoptimised conditions using Lightcycler-480 PCR technology (Roche Diagnostics, Lewes, UK). Specific primers for Notch-1 (Hs00413187_ml), Hrt-1 (Hs01114113_ml), Hrt-2 (Hs00232622_ml) and DLL-4 (Hs00184092_ml) were used and primers for 18S (Hs99999901_sl) ribosomal RNA were used as an endogenous control (Applied Biosystems, Cheshire, UK).
Invasion assay
HMVEC invasion was assayed in Transwell Matrigel Invasion Chambers (BD Biosciences, Oxford, UK). HMVEC (3×104 cells) transiently transfected with Notch-1 siRNA or scrambled control siRNA were added to invasion chambers on precoated matrigel membranes containing EBM supplemented with 2.5% FCS. EBM containing 5% FCS was placed in the lower wells of the assay. Cells were stimulated with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone and in combination for 16 h. Invasion quantification was assessed as previously described.26
Tube formation and cell migration assays
Matrigel (50 µl) (Becton Dickinson, Franklin Lakes, New Jersey, USA) was added to 96-well culture plates and allowed to polymerise at 37°C for 1 h before plating the cells. HMVEC (1×104 cells) transiently transfected with Notch-1 siRNA or scrambled Notch-1 control siRNA were then plated in 250 µl EBM/well onto the surface of the matrigel and stimulated with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone and in combination for 24 h. Furthermore to examine the effect of the synovial microenvironment on EC function, HMVEC were cultured with conditioned media from stimulated synovial explant cultures VEGF, Ang2 alone and in combination±DAPT for 24 h. HMVEC tubes formation was quantified as previously described.26 Wound scratch migration assays were also performed as previously described.54
Cytokine measurements
Interleukin (IL)-6 and IL-8 expression in RA explant cultured supernatants was measured by ELISA according to the manufacturer's instructions (R&D systems).
Activities of MMP-9 and MMP-2 by zymography
MMP-2 and MMP-9 expression in synovial explants and HMVEC supernatants were measured using gelatin zymography (6 mg/ml). 10 µl of supernatants were loaded onto 7.5% polyacrylamide gels. Following electrophoresis, gels were washed with 2.5% Triton X-100 and incubated with substrate buffer at 37°C for 24 h prior to Coomassie brilliant blue stain and destained with water.
Statistical analysis
SPSS V.15 system for windows was used for statistical analysis. For comparisons of multiple groups analysis of variance (ANOVA) with Dunnett post-test for parametric data or the Friedman test with Dunns post-test for non-parametric data was performed. Non-parametric Wilcoxon Signed Rank test for related samples or Parametric Student t tests were performed as appropriate for comparison of two experimental conditions. p<0.05 were considered significant.
Results
Upregulation of Notch signalling by VEGF and Ang2
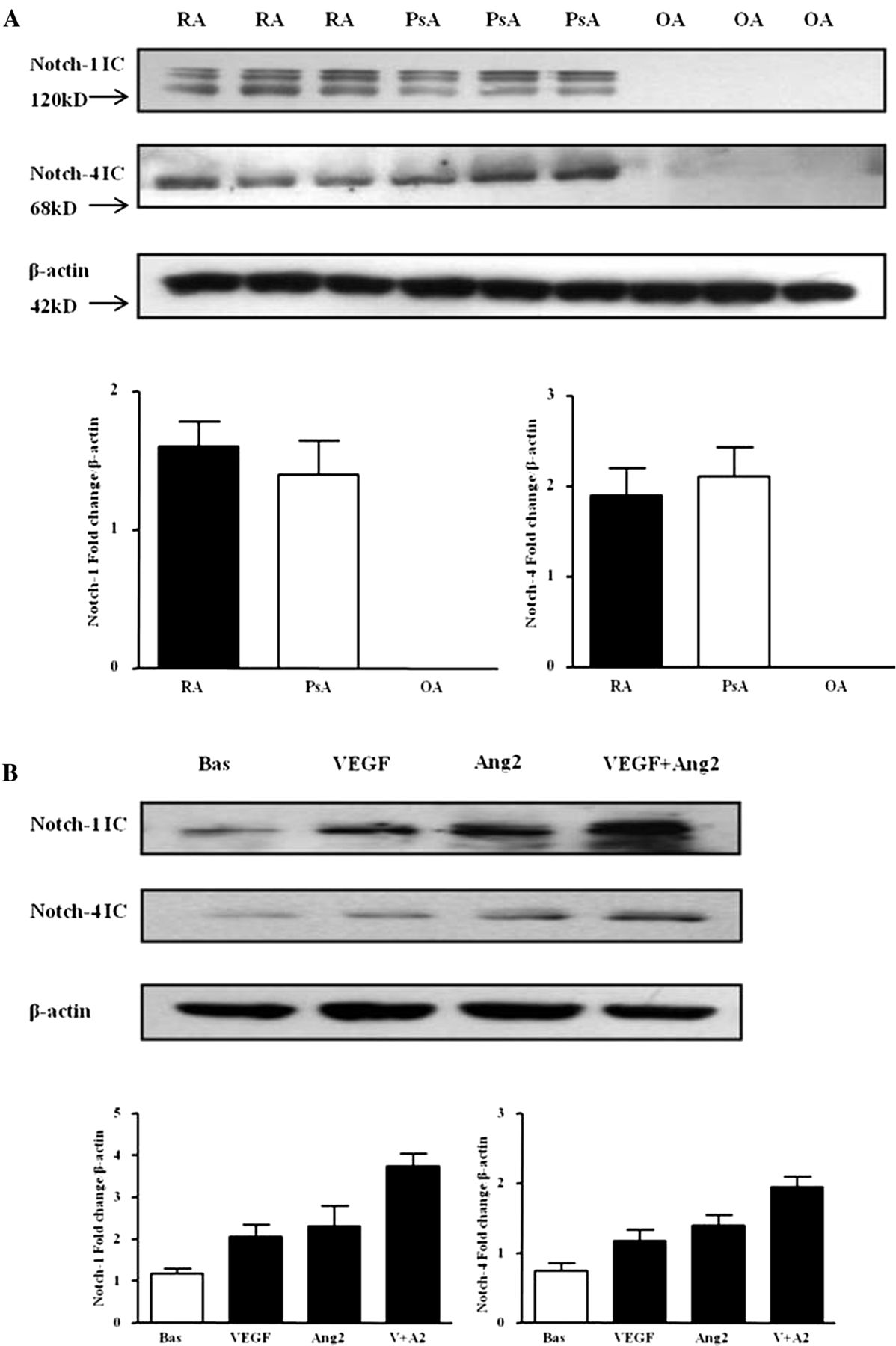
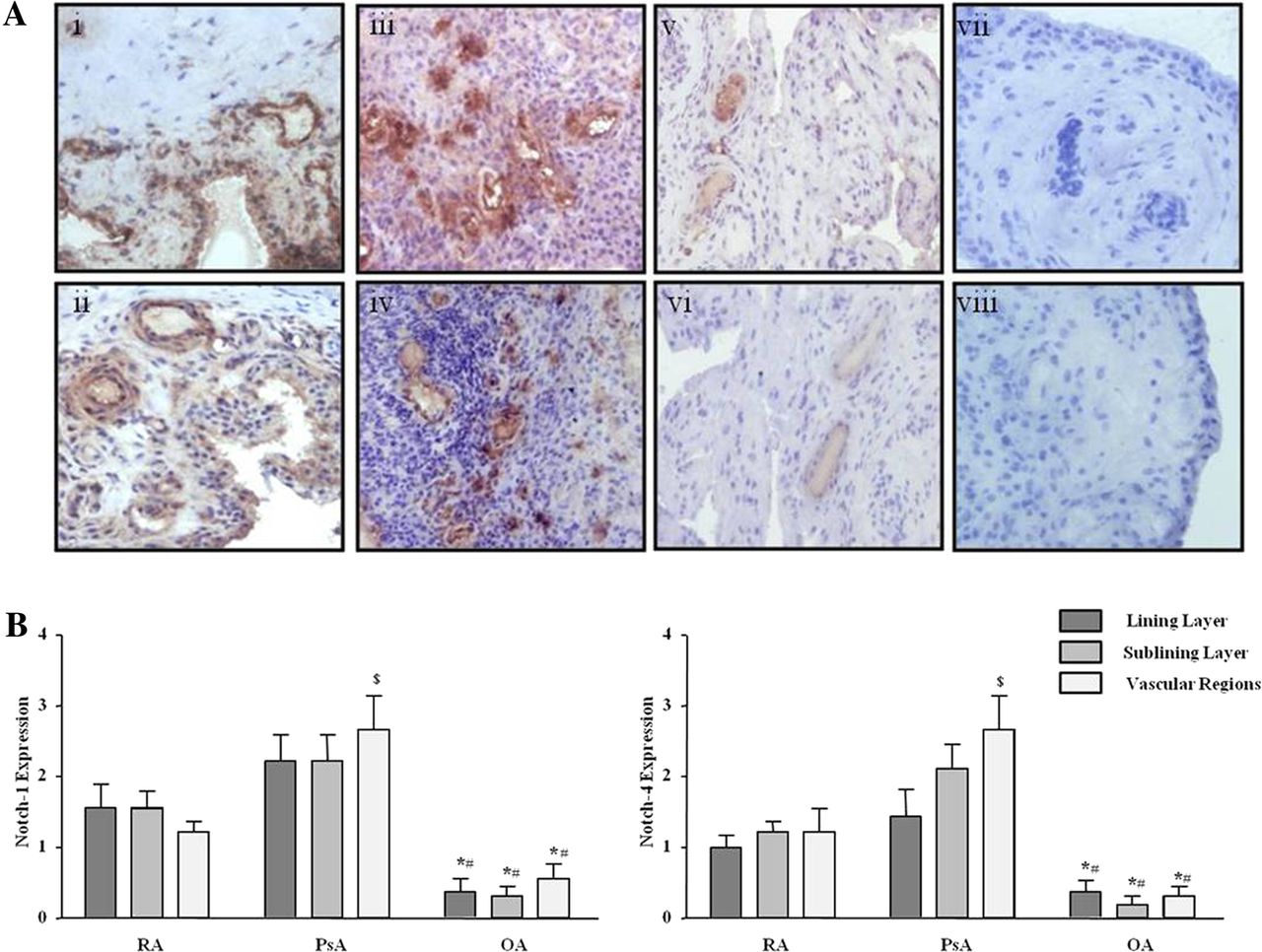
Protein expression of Notch-1 IC (120 kD) and Notch-4 IC (68 kD) were detected in RA and PsA tissue lysates, with minimal expression observed for OA tissue (figure 1A; online supplementary figure S1). Notch-1 IC and Notch-4 IC expression was demonstrated in the synovial perivascular/vascular, sublining layer and lining layer regions in RA (figure 2Ai, ii) and PsA tissue (figure 2Aiii, iv), with minimal expression observed in OA (figure 2Av, vi). No expression for IgG control was observed (figure 2Avii, viii). Semiquantification demonstrated a significant increase in Notch-1 IC and Notch-4 IC expression in RA and PsA patients compared with OA (p<0.05) (figure 2B). Furthermore Notch-1 IC and Notch-4 IC expression in the vascular regions was significantly higher in PsA compared with RA (p<0.05) (figure 2B).
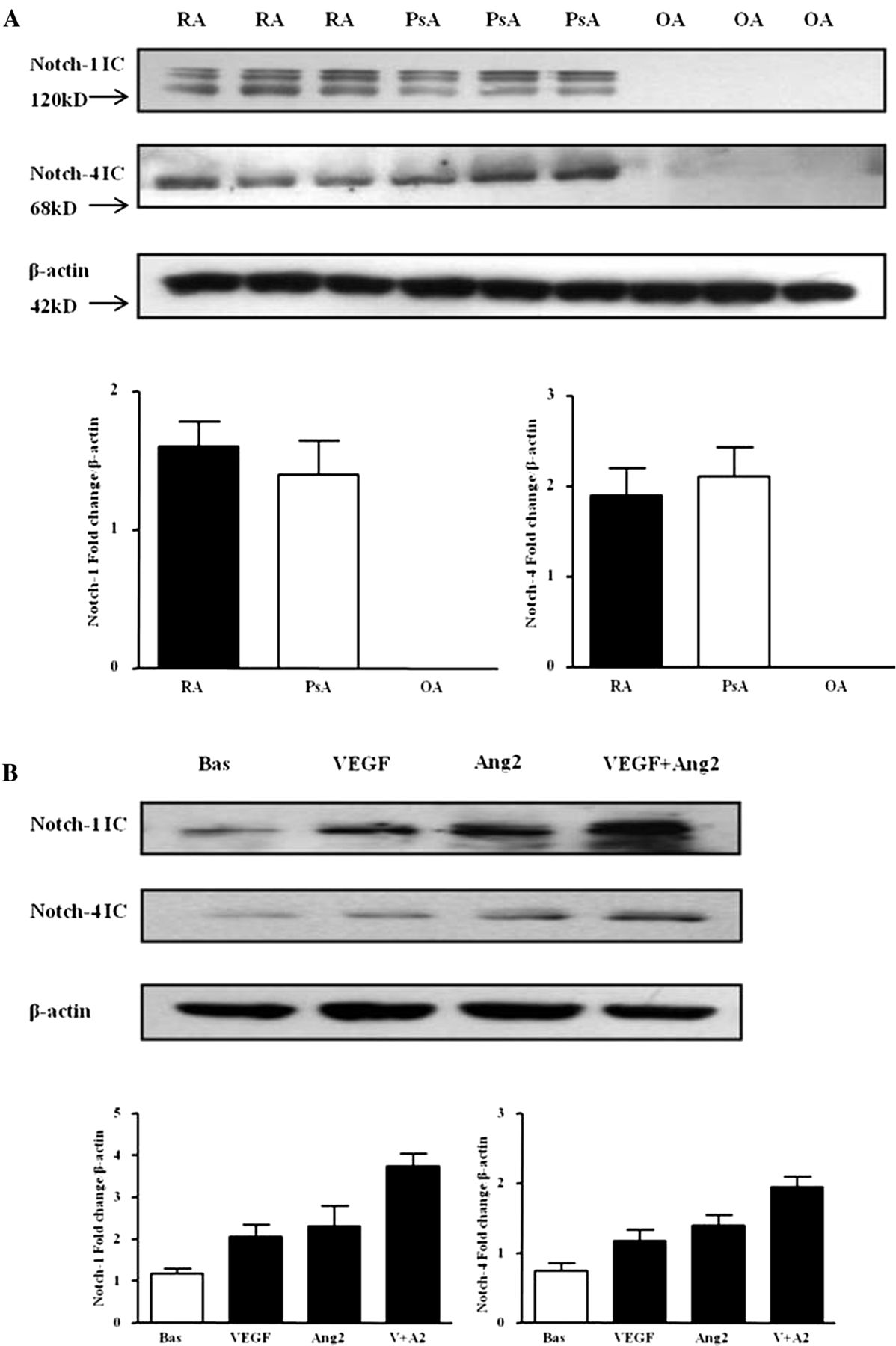
Vascular endothelial growth factor (VEGF) and angiopoietin 2 (Ang2) regulate Notch protein expression in synovial explant cultures. (A) Notch-1 intracellular domain (Notch-1 IC) and Notch-4 IC protein expression were analysed using Western blot in synovial tissues biopsies obtained from RA (n=7), psoriatic arthritis (n=7) and OA (n=6) patients. β-actin was used as loading control. (B) Notch-1 IC and Notch-4 IC protein expression was examined in ex vivo RA synovial explants cultures following stimulation with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone or in combination for 24 h. β-actin was used as loading control. Representative blot of n=3 experiments performed in triplicate.

Notch-1 and Notch-4 localised expression in RA, psoriatic arthritis (PsA) and OA patients. (A) Representative photomicrographs showing Notch-1 and Notch-4 localised expression in synovial tissue from RA (n=10), PsA (n=10) and OA (n=9) patients. Notch-1 and Notch-4 expression are shown in RA tissue (i, ii), PsA (iii, iv), OA (v, vi) respectively. Negative staining for IgG control (vii, viii). (B) Quantification of Notch-1 and Notch-4 in lining, sublining and perivascular regions of RA, PsA and OA tissue. *p<0.05 OA significantly different from RA, #p<0.05 OA significantly different from PsA, $p<0.05 PsA significantly different from RA. Original magnification ×10.
VEGF and Ang2 alone induced Notch-1 IC and Notch-4 IC protein expression, while VEGF/Ang2 combination further increased their expression compared with either VEGF or Ang2 alone (figure 1B). A similar response was observed for PsA explants (see online supplementary figure S2A).
In HMVEC, VEGF and Ang2 alone and in combination induced Notch-1 IC and Notch-4 IC protein expression (figure 3A). Gene expression of Notch-1 receptor, downstream target genes Hrt-1, Hrt-2 and ligand DLL-4 were assessed at 3, 6 and 24 h, with significant induction observed only at 24 h (figure 3B). VEGF and Ang2 stimulation alone significantly induced DLL-4 and Hrt-2 mRNA (p<0.05), with the combination of VEGF/Ang2 further potentiating the expression of Notch-1, its ligand DLL-4 and Hrt-2 (all p<0.05). No significant effect was observed for Hrt-1 (figure 3B).

Vascular endothelial growth factor (VEGF) and angiopoietin 2 (Ang2) regulate the Notch signalling pathway in human microvascular endothelial cell (HMVEC). (A) Notch-1 intracellular domain (Notch-1 IC) and Notch-4 IC protein expression was examined in HMVEC following stimulation with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone or in combination for 24 h by Western blot. β-actin was used as loading control. Representative blot of n=4 experiments. (B) Notch-1, Hrt-1. Hrt-2 and DLL-4 mRNA expression was quantified in HMVEC following stimulation with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone or in combination for 24 h and normalised to Glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Data is representative of three independent experiments. Values are expressed as mean±SEM of n=3 experiments. $p<0.05 significantly different for comparison of multiple parameters by ANOVA, *p<0.05 significantly different from basal, #p<0.05 significantly different from VEGF.
Notch-1 mediates VEGF/Ang2 induced EC invasion and tube formation
Notch-1 siRNA (siN1) transfection resulted in minimal detectable levels of Notch-1 IC compared with scramble control (see online supplementary figure S2B) and dramatically inhibited VEGF-induced expression (see online supplementary figure S2C). Furthermore Notch-1 IC protein expression in HMVEC was significantly decreased in the presence of DAPT (see online supplementary figure S2D).
Figure 4A,B shows representative images and quantification of HMVEC invasion demonstrating significant EC invasion in response to VEGF and Ang2 alone, which was further potentiated by VEGF/Ang2 combination. This effect was significantly inhibited in the presence of Notch-1 siRNA, with no effect for scrambled control (figure 4A,B).

Vascular endothelial growth factor (VEGF) and angiopoietin 2 (Ang2)-induced cell invasion is dependent on Notch signalling. (A) Representative photomicrograph shows human microvascular endothelial cell (HMVEC) invasion under basal conditions (left panel), following VEGF/Ang2 stimulation (middle panel) and in the presence of Notch-1 siRNA (siN1, right panel). At 24 h invading cells attached to lower membrane were fixed (1% glutaraldehyde) and stained (0.1% crystal violet) (hpf, magnification × 40). (B) Representative graphs quantifying non-transfected (control), transfected (Notch-1 or Scramble siRNA), HMVEC invasion following stimulation with VEGF (20 ng/ml), and Ang2 (250 ng/ml) alone or in combination for 16 h. Data are expressed as mean±SEM of replicate experiments (n=4). $p<0.05 significantly different for comparison of multiple parameters by ANOVA, #p<0.05 significantly different from VEGF, *p<0.05 significantly different.
VEGF and Ang2 alone and in combination induced angiogenic tube formation, evident by an increase in the number of EC connecting branches (figure 5A). Notch-1 siRNA transfected HMVEC significantly inhibited VEGF/Ang2 induced tube formation, with no effect for scramble siRNA control. Similar effects were observed for HMVEC migration where DAPT inhibited VEGF/Ang2 combination induced EC migration (see online supplementary figure S3).

Notch signalling mediates vascular endothelial growth factor (VEGF) and angiopoietin 2 (Ang2)-induced cell tube formation. (A) Representative photomicrographs showing human microvascular endothelial cell (HMVEC) tube formation at baseline (left panel), following stimulation with VEGF and Ang2 (middle panel), and in the presence of Notch-1 siRNA transfection (right panel). Quantification of HMVEC tube formation following transfection with Notch-1 or Scramble siRNA following stimulation with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone or in combination. Data are expressed as mean±SEM of replicate experiments (n=4). (B) Representative photomicrograph shows HMVEC tube formation in basal conditioned media containing DMSO as vehicle control (left panel), conditioned media containing VEGF/Ang2 combination (middle panel), and in the presence of 50 µM N-(N-(3,5-Difluorophenacetyl-L-alanly))-S-phenylglycine-t-Butyl Ester (DAPT) (right panel). Connecting branches numbers were quantified following stimulation with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone or in combination. Data are expressed as mean±SEM of replicate experiments (n=4). $p<0.05 significantly different for comparison of multiple parameters by ANOVA or by Friedman analysis, #p<0.05 significantly different from VEGF, *p<0.05 significantly different.
Notch signalling mediates VEGF/Ang2 induced angiogenesis and pro-inflammatory cytokine production in RA explants cultures
To assess the effect of the RA synovial microenvironment on EC function, synovial explants were stimulated with VEGF/Ang2±DAPT. RA synovial explant conditioned media treated with VEGF/Ang2 alone and in combination significantly increased HMVEC tube formation compared with control. This effect was markedly decreased in the presence of DAPT treated explants (figure 5B).
VEGF and Ang2 alone and in combination induced pro-MMP-2 and MMP-9 in RA synovial explants and HMVEC, an effect inhibited by DAPT (figure 6A). Finally, VEGF and Ang2 alone significantly induced IL-6 and IL-8 expression from RA synovial explants with the VEGF/Ang2 combination further potentiated their expression. This effect was significantly inhibited by DAPT (figure 6B). DAPT also inhibited basal cytokine secretion, suggesting that it may effect other pathways that regulate endogenous cytokine secretion.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
N-(N-(3,5-Difluorophenacetyl-L-alanly))-S-phenylglycine-t-Butyl Ester (DAPT) inhibits vascular endothelial growth factor (VEGF)/angiopoietin 2 (Ang2) induced MMPs and cytokines activities. (A) Representative in-gel zymography of pro-MMP-9 and MMP-2 activities (inverted for clarity) in human microvascular endothelial cells (HMVEC) and synovial explants following stimulation with VEGF (20 ng/ml) and Ang2 (250 ng/ml) alone or in combination in the presence of DAPT (50 µM) or DMSO vehicle control. (B) Levels of IL-6 and IL-8 were assayed by ELISA in conditioned media with DAPT (50 µM). Values are expressed as mean±SEM of replicate experiments (n=5). $p<0.05 significantly different for comparison of multiple parameters by ANOVA or by Friedman analysis, #p<0.05 significantly different from VEGF, *p<0.05 significantly different.
Discussion
Angiogenesis and cell migration are critical in the pathogenesis of inflammatory diseases. Here, we demonstrate increased Notch-1 IC and Notch-4 IC protein expression in inflamed synovial tissue compared to OA tissue, and demonstrate that expression is further enhanced by VEGF/Ang2 stimulation of RA/PsA synovial explant cultures and HMVEC. Notch-1 receptor mRNA levels, downstream target genes Hrt-2 and ligand DLL-4 were significantly increased by VEGF/Ang2 stimulation, alone and in combination. In RA synovial explants and HMVEC cultures, VEGF/Ang2 induced EC invasion, angiogenic tube formation, MMPs and cytokine expression, effects that were inhibited by Notch blockade. These data suggest that Notch signalling pathways mediate the pro-angiogenic effects of VEGF/Ang2 in IA.
Angiogenesis occurs early in the pathogenesis of inflammation.1–4 Distinct vascular morphology observed in IA or psoriasis plaques is related to expression of VEGF, Ang and their receptors.5 ,8 ,11 ,12 Cytokines and toll-like receptor (TLRs) induce VEGF, Tie2, Ang1 and Ang2 in synovial fibroblasts and explant cultures.3 ,7 ,26 ,55–58 VEGF mediates hypoxia-induced pro-angiogenic pathways in the inflamed joint.59 ,60 Furthermore, in a CIA model, blocking Tie2 activation inhibited arthritis-induced angiogenesis, receptor activator of nuclear factor Kappa-B ligand (RANKL) expression and bone erosion.25 ,61 This process is dynamic, with studies demonstrating that it is the interactions between the VEGF and Ang-Tie2 pathways that are critical for new vessel formation, stability and survival. In the current study we showed increased expression of Notch-1 IC and Notch-4IC in inflamed synovium compared with OA, with highest expression observed in PsA vascular regions. Furthermore we demonstrated complementary interactions between VEGF and Ang2 acting via the Notch-1 signalling pathway to enhance EC invasion and tube formation, supporting the hypothesis of an unstable synovial vascular microenvironment.3 ,59 ,60 ,62 The previously observed abnormal synovial vascular morphology, differential maturation status and coexpression of VEGF/Ang2 on synovial vessels, suggest that the dysfunctional synovial vessels may be regulated by VEGF/Ang2.5 ,60 ,62 This is consistent with previous reports in cancer models demonstrating that Ang2 blocks vessel stabilisation and induces capillary remodelling and new vessel sprouting in the presence of abundant VEGF.18–21 Furthermore, Hashizume et al, demonstrated that inhibition of Ang2 and VEGF together slows tumour growth by inhibiting sprouting angiogenesis and proliferation.63
In this study, we demonstrate that VEGF and Ang2 alone increase Notch-1 IC and Notch-4 IC protein expression, downstream target genes and the ligand DLL-4, with the VEGF/Ang2 combination potentiating this effect further. In addition, VEGF/Ang2 induced EC invasion, angiogenic tube formation and migration were inhibited by Notch-1 siRNA or DAPT. Furthermore, conditioned media from stimulated RA synovial explants induced EC tube formation, an effect inhibited by Notch blockade. Finally, we demonstrated that VEGF/Ang2 induced cytokine and matrix metalloproteinase (MMP) production was inhibited by DAPT in RA synovial explants, further suggesting Notch mediates VEGF/Ang2 pro-angiogenic effects in the inflamed synovial microenvironment. While the complementary functional interactions of VEGF/Ang2 in cancer models have been described and that VEGF can regulate Notch, this is the first study to show that Notch mediates the downstream functional effects of VEGF/Ang2 interactions in RA explants and HMVEC. In the conditioned media tube formation assays it is unclear which of the growth factors has a stronger effect, as this model maintains the architecture and cell-cell contact of the multiple cells in the synovial tissue, thus other downstream factors may also contribute to the observed functional effect. Previous studies have shown that the relative contribution of VEGF and Ang2 to their complementary action varies depending on the cell type and microenvironment, with studies showing complementary and inhibitory effects of Ang2 on VEGF, 19 ,63 ,64 mechanisms which can also be dependent on the different vascular beds.65
Induction of DLL-4 expression in endothelial tip cells of sprouting vessels in response to VEGF resulting in activation of Notch in adjacent stalk cells has been demonstrated.66 Ang1/Tie2 signalling can induce Notch/DLL-4 through acutely transforming retrovirus (AKT8) in rodent T cell lymphoma-mediated activation of β-catenin.67 Notch-3 also cooperates with hypoxia-inducible factor (HIF)1α to regulate Ang2 expression and neovascularisation under hypoxic conditions.68 Moreover, Notch-1 or Notch-4 mediate cyclic strain-induced angiogenesis and Ang1/Tie2 signalling,69 Consistent with the dysfunctional vascular morphology observed in inflamed joints, differential interactions of Notch with specific ligands has been shown to result in abnormal vessel branching, stability and morphology.13 Moreover, Notch signalling promotes intergrin/focal adhesion kinase (FAK) mediated pathways, which are known to be overexpressed in the inflamed synovium70 ,71 and promote EC branching/sprouting, vacuolation and lumen formation.72–76
Finally, we demonstrate that Notch mediates VEGF/Ang2 induced IL-6 and IL-8 in synovial explants. The observed inhibitory effect of DAPT on basal cytokine secretion also suggests that Notch-1 mediates other pathways involved in regulating endogenous cytokine secretion. This is supported by studies showing Notch signalling can regulate T cell differentiation in RA patients and mediate cytokine production from RA synovial fibroblasts and T helper cells.44 ,46 Notch inhibition downregulates Th17 and Th1 responses in a Giant Cell Arthritis model.77 Furthermore DAPT can inhibit cytokine/chemokine expression from dendritic cells (DC) and T cells78 and can inhibit IL-6 secretion from RA fibroblast-like synoviocytes (FLS).79 Thus, in addition to regulating vascular morphology, Notch signalling may also mediate VEGF/Ang2 induced cytokine profiles in the joint, further exacerbating the inflammatory response and joint destruction.
Beneficial effects of Notch signalling blockade include inducing tumour cell apoptosis, suppressing neuroblastoma cell growth and differentiation,80–82 In contrast, this therapeutic approach may conversely accelerate tumour metastasis and invasion,83 ,84 suggesting that vessel stabilisation, normalisation and improvement of oxygenation are key determinants of outcome.85 Studies also demonstrate Notch signalling is engaged in collagen II (CII)-specific Th1-type and Th17-type expansion, and suggest selective inhibition of Notch signalling pathways as potential therapeutic strategies for the treatment of RA.46
In conclusion, this is the first study to show that VEGF/Ang2 induced pro-angiogenic/inflammatory mechanisms are mediated by Notch signalling pathways in the inflamed joint, further supporting the concept of developing therapeutic targets to the VEGF/Ang2-Notch axis in the treatment of IA.
Acknowledgments
This work was supported by the Health Research Board, Ireland.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
Footnotes
-
Contributors All authors contributed to this manuscript. UF had full access to all of the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis. Study conception and design; Acquisition of data; Analysis and interpretation of data: WG, CS, CW, PR, JMC, DJV, UF.
-
Funding Health Research Board, Ireland.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval This study was conducted with the approval of the St. Vincent's University Hospital ethics committee.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Open Access This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 3.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/3.0/