Article Text

Abstract
Objectives To establish the role and effect of glucocorticoids and the endogenous annexin A1 (AnxA1) pathway in inflammatory arthritis.
Methods Ankle joint mRNA and protein expression of AnxA1 and its receptors were analysed in naive and arthritic mice by real-time PCR and immunohistochemistry. Inflammatory arthritis was induced with the K/BxN arthritogenic serum in AnxA1+/+ and AnxA1−/− mice; in some experiments, animals were treated with dexamethasone (Dex) or with human recombinant AnxA1 or a protease-resistant mutant (termed SuperAnxA1). Readouts were arthritic score, disease incidence, paw oedema and histopathology, together with pro-inflammatory gene expression.
Results All elements of the AnxA1 pathway could be detected in naive joints, with augmentation during ongoing disease, due to the infiltration of immune cells. No difference in arthritis intensity of profile could be observed between AnxA1+/+ and AnxA1−/− mice. Treatment of mice with Dex (10 µg intraperitoneally daily from day 2) afforded potent antiarthritic effects highly attenuated in the knockouts: macroscopic changes were mirrored by histopathological findings and pro-inflammatory gene (eg, Nos2) expression. Presence of proteinase 3 mRNA in the arthritic joints led the authors to test AnxA1 and the mutant SuperAnxA1 (1 µg intraperitoneally daily in both cases from day 2), with the latter one being able to accelerate the resolving phase of the disease.
Conclusion AnxA1 is an endogenous determinant for the therapeutic efficacy of Dex in inflammatory arthritis. Such an effect can be partially mimicked by application of SuperAnxA1 which may represent the starting point for novel antiarthritic therapeutic strategies.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits use, distribution, and reproduction in any medium, provided the original work is properly cited, the use is non commercial and is otherwise in compliance with the license. See: http://creativecommons.org/licenses/by-nc/3.0/ and http://creativecommons.org/licenses/by-nc/3.0/legalcode
Statistics from Altmetric.com
Introduction
There is genuine interest in detailing cellular and molecular events regulating the phase of the resolution of inflammation, with the characterisation of specific players and targets.1 This interest stems from the appreciation that, in ideal settings, a strong pro-inflammatory response, orchestrated by cytokines, adhesion molecules and chemoattractants2 must be followed by an anti-inflammatory and pro-resolving phase, which assures spatial and timely control of the response the host organises upon encounter with an insult,3 leading to resolution and regain of tissue functions.
Annexin A1 (AnxA1) is an effector of resolution.4 Highly expressed in immune cells (eg, polymorphonuclear cells and macrophages), this protein is externalised to exert paracrine and juxtacrine effects, the vast majority of which are mediated by the formyl-peptide receptor type 2 (FPR2/ALX ([Lipoxin A4 receptor]) or FPR2, in rodents).5 Intriguingly, FPR2/ALX is also the lipoxin A4 receptor6 indicating the existence of important – yet not fully appreciated – networks in resolution.7 Another receptor is also advocated to mediate the effects of AnxA1, the formyl-peptide receptor type 1 or FPR1 (FPR1 in rodents),8 though it is unclear if and how this receptor binds the full-length protein9 or whether – more selectively – it would mediate effects of the AnxA1 peptidomimetic peptide Ac2-26.10
The majority of experimental approaches used to define properties of pro-resolving mediators have been reliant on models of acute inflammation, characterised by rapid polymorphonuclear influx followed by inflammatory monocytes which differentiate into tissue macrophages3 ,11 to phagocytose debris and apoptotic cells and favour tissue repair, a conclusive act in resolution that may require re-epitheliation12 or a shift in fibroblast phenotype.13 In this context, the pro-resolving nature of AnxA1 has been demonstrated in a variety of experimental settings, noting induction of apoptosis, promotion of efferocytosis and tissue repair, spanning from models of acute inflammation (eg, 14 ,15) to a model of colitis where delayed resolution was observed in AnxA1−/− mice.16 An important aspect of AnxA1 biology is its second messenger role in the actions of glucocorticoids, a function mainly addressed in cellular settings (eg, 17) or in models of acute inflammation.18 ,19 However, Yang's group applied antigen-induced arthritis to observe reduction afforded by dexamethasone (Dex) on synovitis, soft tissue inflammation and cartilage erosion attenuated in AnxA1−/− mice.20
The K/BxN serum-induced model of inflammatory arthritis is ideal for investigating the impact of specific pro-resolving pathways. Using this model of active phase of rheumatoid arthritis, Krönke et al reported a higher degree of joint inflammation in 12/15-lipoxygenase−/− mice, associated with their inability to synthesise lipoxin A4.21 Similarly, animals nullified for pro-resolving melanocortin receptor type 3 displayed higher degree of arthritis and delayed resolution, accompanied by augmented cytokine expression and osteoclast responsiveness.22 An equally interesting study investigated the effect of absence of glucocorticoid signal in osteoblasts, by means of restricted transgenic expression of 11β-hydroxysteroid dehydrogenase type 2, in this model of inflammatory arthritis. The outcome was that of a more attenuated arthritis in the transgenic mice when assessed macroscopically and histologically, and selectively in the delayed phase, after peak disease.23
The present study made use of the serum-induced model of inflammatory arthritis to: (1) monitor potential modulation of endogenous AnxA1, FPR1 and FPR2; (2) determine the effect of Dex in wild type and AnxA1−/− mice; and (3) establish the pharmacological potential of exogenously administered AnxA1.
Materials and methods
Animals
BALB/c male 8-week old wild type AnxA1+/+ and AnxA1−/− mice24 were bred at Charles River (Kent, UK). All experiments were approved and performed under the guidelines of the Ethical Committee for the Use of Animals, Barts and The London School of Medicine and Home Office regulations (Scientific Procedures Act, 1986).
Model of inflammatory arthritis
Arthritis induction
Mice received either 50+50 µl (day 0 and 2) or 200 µl (day 0) of arthritogenic K/BxN serum.22 Disease was monitored by assessing the clinical score where a maximum of 12 points could be given per animal: 0 point, no evidence of inflammation; 1 point, subtle inflammation on metatarsal phalanges joints, individual phalanx; 2 points, swelling on dorsal or ventral surface of paw; 3 points, major swelling on all aspects of paw.25 Paw volume was assessed by water plethysmometry (Ugo Basile, Milan, Italy).
Pharmacological intervention
Mice were injected intraperitoneally with Dex (disodium salt; Sigma, Dorset, UK) prepared in sterile phosphate buffered saline (Sigma) and given at a dose range of 3–30 µg once daily after disease onset (from day 2). AnxA1 and SuperAnxA1 were generated as recently described26 and administered intraperitoneally, from day 2, at 1 µg per mouse.26
Histology and immunohistochemistry
Histology
Joints were decalcified and paraffin embedded. Sections (4 micrometers) were stained with H&E or safraninO with fast green counterstain. Standard light microscopy was used to determine the degree of synovitis, pannus formation, cartilage and bone erosion which were graded from 0 (no disease) to 5 (severe) by two blinded examiners, as reported.22
Immunohistochemistry
An antigen retrieval step using citrate buffer (pH 6.0) was performed and then sections were incubated with rabbit polyclonal anti-AnxA1 (1:1000) (Zymed Laboratories, Cambridge, UK) or anti-FPR2 (1:4000).27 After washing and the secondary biotinylated antibody (Dako, Cambridge, UK), staining was detected using a peroxidase conjugated streptavidin complex (Dako) and colour developed using 3,3' Diaminobenzidine (DAB) substrate (Sigma).
Molecular analyses
Ankle joints were snap frozen in liquid nitrogen and homogenised in Qiagen RLT plus buffer (Qiagen, Crawley, UK) using Precellys24 ceramic bead homogenisation technology (Bertin Technologies, Montigny-le-Bretonneux, France). RNA was extracted using RNeasy Plus mini kit (Qiagen) and genomic DNA contamination eliminated with Turbo DNA-free kit (Applied Biosystems, Foster City, California, USA). Complementary DNA was synthesised using SuperScriptIII reverse transcriptase and OligoDt primers (Invitrogen). Quantitative real-time PCR was performed using QuantiTect primers (Qiagen) and ABI Prism 7900 sequence detector system (Applied Biosystems). RQ values were calculated using the 2−( ΔΔCt) method and normalised to an individual naive AnxA1+/+ or AnxA1−/− mouse (calibrator sample).
All Ct values were normalised to endogenous Gapdh gene product. Since factors such as hypoxia can influence Gapdh gene levels,28 a second house-keeping gene Rpl32 was used to normalise and confirm data (see table S2).
Statistics
Data are expressed as mean±SE. Student t test was used to compare two groups. Comparison of clinical scores and paw volumes were made using 2-way analysis of variance. A value of p<0.05 was considered significant.
Results
Characterisation of joint inflammation
We began this study to optimise the model by comparing two different protocols: 200 µl of serum at day 0 (as in22) or 50+50 µl at day 0 and day 2. A remarkably similar arthritic response could be measured with respect to disease profile and intensity (figure 1A). High levels of disease were achieved as early as day 2 postinjection with maximal response by day 6, after which the disease resolved to almost normal by day 20. Incidence of disease was similar between the two volumes of K/BxN serum (table S1). The arthritic response was mirrored by changes in paw volume with similar profiles with no difference between the two protocols (figure 1B).

Characterisation of the arthritic response to low volume K/BxN serum. Mice (n=8) were given an intraperitoneal injection of 50 μl at day 0 and day 2 (50+50 μl) or a single injection of 200 μl at day 0 of K/BxN serum. (A) Arthritic scores and (B) paw oedema were monitored over the time course. No significant difference (two-way analysis of variance) was observed between the two K/BxN serum groups. (C) Ankles of naive, 50+50 μl and 200 μl K/BxN serum group. Decalcified paraffin embedded joints were stained by H&E and safraninO. Representative images are shown; scale bars, 50 μm. CE, cartilage erosion; PF, pannus formation; S, synovitis. (D) Histomorphometric analyses of joint sections. All results are expressed as mean±SE.
Microscopic analyses of non-arthritic ankle joints showed an acellular articular cavity with intact unicellular synovial lining and uniform cartilage structure (figure 1C, top panel). In contrast, arthritic joints from both 50+50 µl and 200 µl K/BxN arthritis groups displayed marked cell infiltration within a thickened synovial lining. Pannus invasion leading to cartilage erosion was observed and proteoglycan loss was confirmed with safraninO (figure 1C, middle and bottom panels). Quantitative histological scoring showed no significant difference between the two groups of arthritic mice (figure 1D).
The ankle is the predominant joint to be affected in this model of inflammatory arthritis, yet we monitored structural changes in the knee, as is perhaps more relevant to human disease; we observed that knee joints displayed mild synovitis accompanied with thickening of synovial lining, pannus formation and marked cartilage erosion (see figure S1). Based on these results, in agreement with a recent published study,29 the 50+50 µl serum protocol was selected for subsequent experimentation.
Expression of AnxA1 and FPR2 in the joint
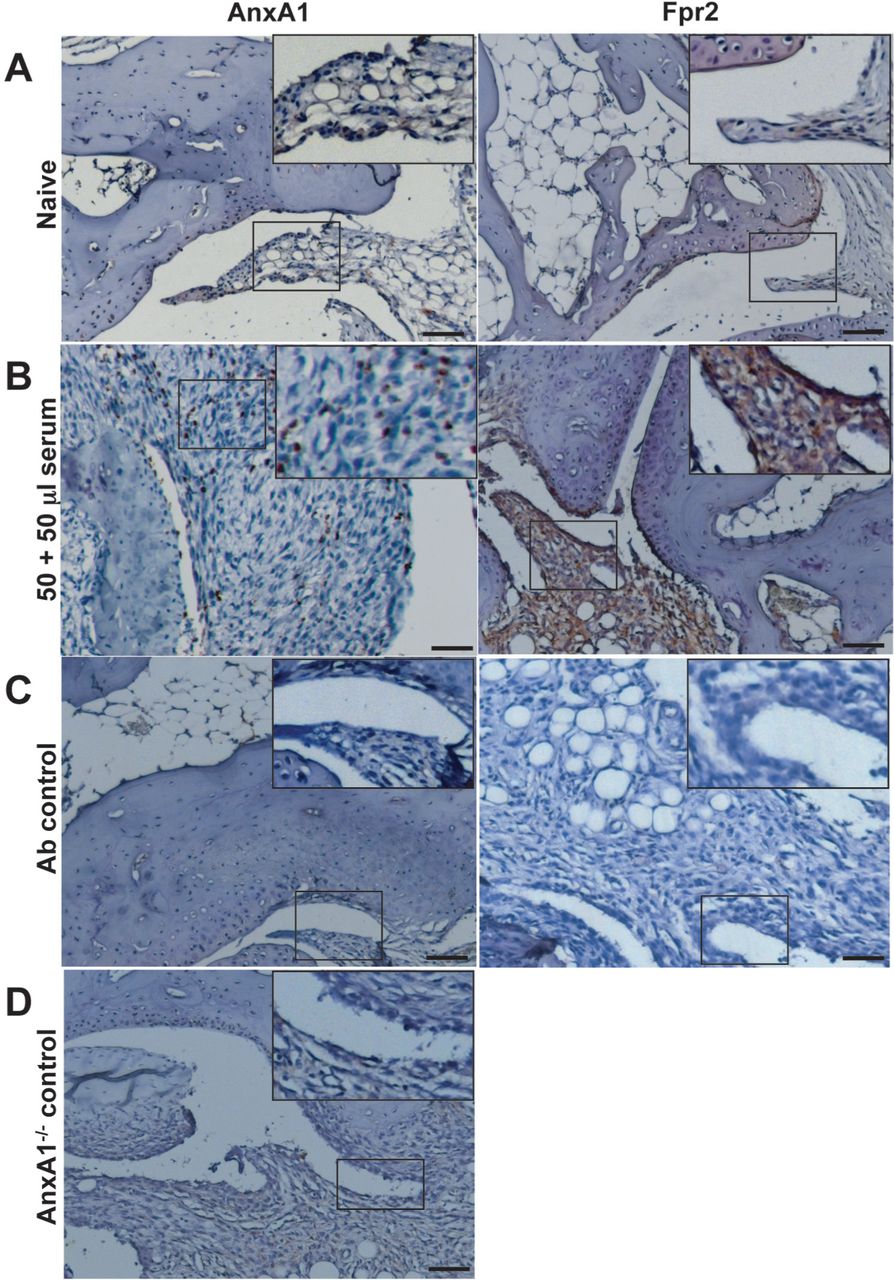
At the mRNA level, a low degree of expression for both FPR1 and FPR2 could be measured, with high AnxA1 message content (see table S2). These results were partially confirmed by protein analyses, since AnxA1 was mildly expressed and localised predominantly in the synovial lining, while FPR2 was localised to the periosteal lining with very little detection in synovium (figure 2A). In arthritic joints, AnxA1 and FPR2 expression was highly detectable within the synovial lining (FPR2) and tissue (AnxA1 and FPR2) (figure 2B). Antibody controls and AnxA1−/− joints did not show any staining (figure 2C,D).

Protein expression of AnxA1 and FPR2 in ankle joints of naive mice. Ankles of naive and K/BxN serum (50+50 μl, day 0 and day 2) were processed (see methods) and stained for AnxA1 and FPR2, with a haematoxylin counterstain. Images representative of three distinct analyses in four mice per status are shown. (A) Naive mouse ankle joints. (B) Serum-treated mouse ankle joint. (C) Same joints but tested without the primary antibody. (D) Joint of an AnxA1−/− mouse acting as negative control for AnxA1 staining. Scale bars, 20 μm.
Dex is inactive in AnxA1−/− mice
Despite wide application of this model of inflammatory arthritis, the bioactions of glucocorticoids have been scantly investigated. We selected the 10-µg dose of Dex from initial studies (figure S2). Administration of Dex to AnxA1+/+ mice exerted profound inhibitory effects with ∼60% inhibition of the severity of arthritis at day 6 and ∼80% at day 10 compared with vehicle-treated mice (figure 3A). AnxA1−/− mice displayed a similar profile of arthritis to AnxA1+/+ animals. However, the antiarthritic effect of Dex was absent in animals nullified for AnxA1 (figure 3A). Analysis of presence of severe disease (score ≥8) indicated a trend for higher incidence in AnxA1−/− mice and a marked reliance on endogenous AnxA1 for the protective effects of Dex (table S3).

Dexamethasone (Dex) attenuates K/BxN arthritis in AnxA1+/+ but not AnxA1−/− mice. AnxA1+/+ and AnxA1−/− mice were given an intraperitoneal injection of K/BxN serum (50 μl at day 0 and day 2) and then received vehicle or Dex (10 µg intraperitoneally daily). (A) Arthritic scores as monitored over a 10-day time course. *p<0.05 versus appropriate vehicle control (two-way analysis of variance). (B) Right ankles of mice from each experimental group were taken at day 10 and joints processed for staining by H&E and safraninO. Representative images are shown. Scale bars, 50 μm. CE, cartilage erosion; PF, pannus formation; S, synovitis. (C) Histomorphometric analyses of joint sections. *p<0.05 versus appropriate vehicle control (Student t test). (D) Quantitative real-time PCR was performed on cDNA from left ankle joints; pro-inflammatory genes were analysed with all Ct values normalised to endogenous Gapdh (four mice per group). RQ values were calculated using 2−(ΔΔCt) and data shown here as mean % gene inhibition by Dex relative to vehicle control groups±SE. Naive joints were set as the calibrator samples. *p<0.05 versus respective vehicle control (Student t test).
Histological analyses were in line with arthritic scores. AnxA1+/+ mice displayed marked synovitis accompanied by pannus formation and cartilage destruction (figure 3B, left panels). Treatment with Dex prevented signs of disease (figure 3B, middle panels), while AnxA1−/− mice treated with vehicle control (not shown) and Dex (figure 3B, right panels) showed similar disease pathology to AnxA1+/+ mice. These structural changes were reflected in the histological scores with ∼75% inhibition afforded by Dex only in AnxA1+/+ mice (figure 3C).
Analyses of inflammatory gene messages, normalised to Gapdh and Rpl32 (see figure S3) and selected from our previous study,22 revealed that in wild type mice Dex inhibited Cxcl10, Nos2, interleukin (IL)-1β and Ccl2. Tumour necrosis factor α (TNFα) was inhibited by ∼25% while expression of the chemokine receptor Cxcr3 was not modulated (figure 3D). These inhibitory effects of Dex were absent or significantly reduced in AnxA1−/− mice.
Next, we monitored expression of AnxA1 and its receptors, normalised to Gapdh and Rpl32 (see figure S4). In arthritic AnxA1+/+ mice, both FPR1 and FPR2 mRNA expression was enhanced over naive joints (figure 4), a finding which is in line with FPR2 protein expression (see figure 2). Dex decreased both FPR1 and FPR2 message with little comparative effect on AnxA1 (figure 4). In mice devoid of AnxA1, a >10-fold increase in FPR1 expression was observed comparing arthritic to naive joints, with FPR2 remaining at a similar expression. In absence of AnxA1, Dex could not regulate either FPR1 or FPR2 gene expression (figure 4).

Profile of AnxA1, FPR1 and FPR2 gene product expression in ankle joints of AnxA1+/+ and AnxA1−/− mice treated with dexamethasone (Dex). AnxA1+/+ and AnxA1−/− mice were given an intraperitoneal injection of K/BxN serum (50 μl at day 0 and day 2) and then received vehicle or Dex (10 µg intraperitoneally daily). Quantitative real-time PCR was performed on cDNA from left ankle joints of mice from each experimental group (day 10). The AnxA1, FPR1 and FPR2 genes were analysed with all Ct values normalised to endogenous Gapdh (four mice per group). Naive joints were set as the calibrator samples. Mean RQ values±SE shown here were calculated using 2−(ΔΔCt) method. *p<0.05 versus naive; §p<0.05 versus appropriate vehicle control; #p<0.05 versus AnxA1+/+ group (Student t test).
SuperAnxA1 accelerates resolution of the arthritic joint
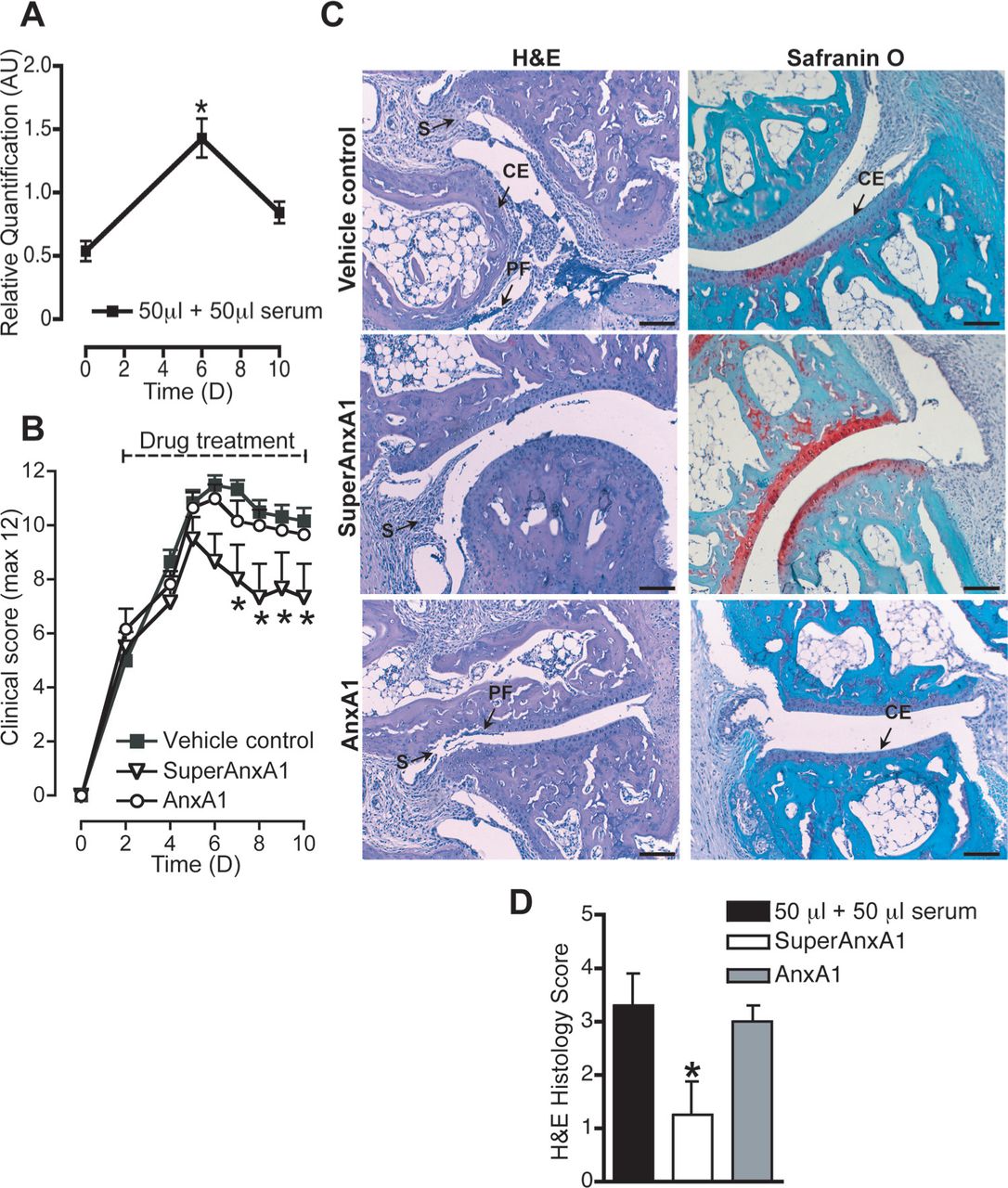
In the final set of experiments, we assessed the pharmacological potential of these findings. Proteinase 3 (PR3) may limit AnxA1 functions in neutrophilic inflammation, cleaving it at the N-terminal region. Thus, we monitored the profile of PR3 gene in the arthritic joint and found it significantly rose from baseline (day 0) to peak of disease (day 6) (figure 5 and figure S5). This observation, coupled with abundant expression of FPR1 and FPR2 in the joints justified the testing of SuperAnxA1. Dosed at 1 µg per mouse, AnxA1 was not particularly effective in reducing disease (figure 5B). On the other hand, cleavage-resistant SuperAnxA1 did not affect disease progress (eg, from day 2 to day 6) but significantly accelerated resolution (figure 5B), with 50% less disease incidence compared with controls (table 1).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
SuperAnxA1 accelerates resolution of K/BxN arthritis. AnxA1+/+ mice (n=6) were given an intraperitoneal injection of K/BxN serum (50 μl at day 0 and day 2) and then received vehicle, human recombinant AnxA1 or SuperAnxA1 (1 µg intraperitoneally daily from day 2). (A) PR3 gene product expression was analysed with Ct values normalised to endogenous Gapdh. RQ values were calculated using 2−(ΔΔCt) and data shown here as mean±SE. Naive mice (D0) were set as the calibrator samples. *p<0.05 versus naive (Student t test). (B) Arthritic score. *p<0.05 versus vehicle control (two-way analysis of variance). (C) Right ankles of mice from each experimental group were taken at day 10 and joints processed for staining by H&E and safraninO. Representative images are shown. Scale bars, 50 μm. CE, cartilage erosion; PF, pannus formation; S, synovitis. (D) Histomorphometric analyses of joint sections. *p<0.05 versus vehicle control (Student t test).
SuperAnxA1 reduces the severity of inflammatory arthritis
The macroscopic joint protection afforded by SuperAnxA1 was confirmed microscopically, with preservation of cartilage structure with little loss of proteoglycans (figure 5C, middle panels). Synovitis was apparent within these joints, but substantially lower than controls (figure 5C, top panels). The joints of AnxA1 treated mice were disrupted to a similar extent as controls (figure 5C, bottom panels). Quantitative analysis of damage within the ankle joints confirmed the observational studies (figure 5D). Finally, ankle joints from vehicle- and SuperAnxA1-treated mice demonstrated reduced expression of the Cxcl10 and Cxcr3 pair, as well as Nos2, IL-1β, Ccl2, IL-6 but not IL-10 and TNFα (figure S6).
Discussion
AnxA1 is the prototype of a new class of mediators able to potently modulate both the innate and adaptive immune response; while effecting inhibitory properties on inflammatory cells, setting in motion pro-resolving responses, this protein acts as a positive relay in T cell activation.4 As an example, exogenous and endogenous AnxA1 potentiate T cell receptor-downstream signalling30 and assure proper dendritic cell-mediated T cell clonal expansion.31 In agreement with these dual functions, AnxA1−/− mice experience augmented acute inflammatory responses,14 ,15 yet display reduced degree of tissue damage in an experimental model of multiple sclerosis.32 Less clear is the role of the protein in models of arthritis.
In the collagen-induced arthritis model, AnxA1 injection during the immune phase (within 1 week postcollagen immunisation) led to higher disease as emerged 3 weeks later.30 In a model of antigen-induced arthritis, assessed at a single time-point (day 7), endogenous AnxA1 modulated some of the anti-inflammatory effects of Dex, including inhibition of pro-inflammatory genes including IL-1β and TNFα.20 No studies have yet investigated the role of endogenous or exogenous AnxA1 in the serum- induced inflammatory arthritis model.
K/BxN serum-induced arthritis33 ,34 is a versatile model that can be applied to transgenic mice with little dependence on strain.35 This model addresses the contribution of neutrophils and macrophages to inflammatory arthritis, as evident from depletion studies.36 ,37 Conversely, this experimental arthritis does not rely on T and B cells as demonstrated with the RAG1−/− mice28 which, in contrast, display delayed disease onset and severity in collagen-induced arthritis.38 Here we describe that low dosage arthritogenic serum provokes a high degree of arthritis and disease incidence, data which are in agreement with a recent study by Boilard's group.29 We noted a remarkable joint disruption and report a novel parallelism between the ankle and knee joint. Marked synovitis associated with disruption of cartilage occurs along a well-defined time-profile with peak disease at day 6 followed by a resolution phase.22 ,35 Use of transgenic mice has allowed some definition of the disease pathogenesis, revealing a fundamental role for myeloid cells36 ,37 and IL-1.39 ,40 More recently, this model of arthritis has been applied to pro-resolving pathways, noting that 12/15-lipoxygenase or melanocortin type-3 receptor deficient mice experience a higher degree of disease.21 ,22
AnxA1 is highly abundant in myeloid and stromal cells and its expression can be modulated by glucocorticoids and other anti-inflammatory drugs.41 In addition, cell activation such as adhesion or extravasation also engages the AnxA1 system as a way to activate an inhibitory circuit to modulate the inflammatory response.27 After several pharmacological investigations with AnxA1 or mimetics, AnxA1 null mice were produced and tested in models of acute inflammation15 ,24 revealing important modulatory functions.4 As discussed above, AnxA1 role in chronic inflammatory settings in less defined.
This study began by detecting protein and mRNA of elements of the AnxA1 pathway during ongoing inflammatory arthritis, reporting a high degree of expression for AnxA1, FPR1 and FPR2 both in resting and arthritic joints. At the protein level, AnxA1 localisation in the mouse synovium seems to parallel that observed in the human rheumatoid arthritis synovium.42 However, at least in the conditions applied here, lack of AnxA1 does not seem to impact on the profile of disease. Yet, it remains to be seen whether such a lack of function would also be seen in knockouts for the AnxA1 receptors.5 ,43 As an example, in preliminary observations, mice nullified for FPR2 displayed higher arthritic score upon injection of the arthritogenic serum.44
Glucocorticoids have been scarcely tested in this model. Buttgereit et al have tested the effect of osteoblast-restricted absence of glucocorticoid signalling on disease outcome, noting a worsening with marked joint disruption.23 In these mice, bone cells could not respond to glucocorticoids from birth; thus, it is possible that developmental aspects might be relevant and subtle long-term alterations occur. For instance, it is known that glucocorticoids exert a permissive effect on important cytokine function in the liver, by assuring proper expression of the IL-6 and IL-1 receptors.45 ,46 IL-6 is also a master-cytokine for osteoblasts (and fibroblasts) and so a malfunctioning or inadequate glucocorticoid signalling, prolonged from birth, may generate responses that are not necessarily replicated by acute glucocorticoid treatment.
In keeping with this, our pharmacological analyses with doses of Dex as low as 10 µg exerted potent protective effects on the joint, evident both as inhibition of pannus formation and chondro-protection. It was striking to observe that the effects of the glucocorticoid were lost in AnxA1−/− animals. Thus, while AnxA1−/− mice did not experience a difference in the extent and profile of arthritis, at variance from the antigen-induced arthritis model,20 endogenous AnxA1 still proved important in the inhibitory effects of Dex on a discrete set of pro-inflammatory genes, of which IL-1β, Cxcl10 and Ccl2 were of particular interest. Selective modulation of gene expression downstream AnxA1 is in keeping with the observations made in the antigen-induced arthritis model.20 The fundamental role that IL-1β exerts in this model of inflammatory arthritis has been addressed through a variety of genetic approaches targeting directly the cytokine, its receptor39 ,47 or endogenous antagonist.40
Modulation of gene expression downstream of endogenous AnxA1 can also be discussed in relation to the modulation of genes of the AnxA1 pathway. In fact, Dex did not appear to induce AnxA1 or FPR gene expression at the level of the joint, yet was highly reliant on endogenous AnxA1 both with respect to cell recruitment and inflammatory gene expression. We reason that these data indicate multiple functions of endogenous AnxA1, which become of relevance only after administration of the glucocorticoid.
The lack of correlation between AnxA1 gene product and protein is intriguing though it should be said that glucocorticoid- regulation of the AnxA1 pathway is multifaceted; thus, non-genomic modulation48 with release of the protein, for instance, could account for apparent lower tissue levels of the AnxA1 protein. AnxA1 analysis, at a single time point, could also be confounded by genomic regulation exerted by glucocorticoids15 ,49 and other factors present in an inflamed joint such as IL-650 and TNF.51 Clearly, the current lack of characterisation of the AnxA1 promoter contributes to this poor understanding of the mechanisms operative in these settings. Moreover, the fact that glucocorticoids like Dex induce AnxA1 gene in innate immune cells14 ,15 while downregulating it in adaptive immune cells52 adds another layer of complexity to this issue which inevitably requires further experimentation.
The versatility of this model of inflammatory arthritis justifies its application to a variety of targets; in line with the major infiltration of neutrophils and mast cells,53 we could observe dynamic expression of PR3 gene product in the arthritic joint. PR3 is emerging as an important effector of neutrophilic inflammation54 and can cleave, among a variety of substrates, also AnxA1.55 Therefore, we concluded this study by testing the potential of AnxA1 mimetics, comparing the effect of native protein with that of SuperAnxA1, a mutant resistant to serine protease-induced cleavage.26 Administered at the dose of 1 µg (corresponding to 27 pmol), SuperAnxA1, but not AnxA1, attenuated joint disease. It was interesting to note that SuperAnxA1 did not impact on the induction of arthritis yet it accelerated the resolution process so that by day 10 a preserved joint structure could be observed. We wish to propose that higher resistance to proteolysis might underlie its pharmacological efficacy, though we cannot exclude the confounding element of potential differences in pharmacokinetics. In any case, these results provide strong proof-of-concept that an AnxA1 derivative, resistant to proteolysis, could be beneficial in modulating joint inflammation. SuperAnxA1 could be the backbone for future strategies aiming at capitalising AnxA1 biology for novel antiarthritic approaches.
Glucocorticoids are of wide clinical use because of their ability to impact several effectors of arthritis, with a recent appreciation that, besides inhibition of pro-inflammatory gene expression, gene induction is also of therapeutic relevance.56 ,57 In the context of chronic inflammatory diseases, induction of dual-specificity phosphatase 158 and glucocorticoid-induced leucine zipper59 genes could be of mechanistic value. In addition, we show here that endogenous AnxA1 is also relevant to the antiarthritic effect of low-dose Dex, yet we cannot demonstrate induction of this gene at the level of the arthritic joint leading to our proposal that the protein is either modulated in a non-genomic fashion by this glucocorticoid41 ,60 or the regulation occurs outside the site of the arthritic joint, an obvious site being the vasculature and the control of the egress of bloodborne neutrophils.4 Analysis of AnxA1 expression in distant tissues (eg, blood) during ongoing arthritis and glucocorticoid treatment can shed light on this point, which has also been touched upon above in relation to lack of correlation between joint AnxA1 mRNA and protein expression.
Collectively, this study reveals potent antiarthritic properties for Dex in a model of inflammatory arthritis, using very low doses perhaps reminiscent of clinical application on low-dose glucocorticoids in RA61 and, in terms of drug discovery programmes, justifies the testing of AnxA1 mimetics to control the active phase of the disease.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Web Only Data - This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
-
Funding This work was supported by a Wellcome Trust (UK) project grant 083551. SMO is funded by Fundação de Amparo à Pesquisa do Estado de São Paulo – FAPESP (Grant 2011/00128-1) and Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq (Grant 302768/2010-6).
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.