Article Text

Abstract
Objectives Spondyloarthritides (SpA) are characterised by both peripheral and axial arthritis. The hallmarks of peripheral SpA are the development of enthesitis, most typically of the Achilles tendon and plantar fascia, and new bone formation. This study was undertaken to unravel the mechanisms leading towards enthesitis and new bone formation in preclinical models of SpA.
Results First, we demonstrated that TNFΔARE mice show typical inflammatory features highly reminiscent of SpA. The first signs of inflammation were found at the entheses. Importantly, enthesitis occurred equally in the presence or absence of mature T and B cells, underscoring the importance of stromal cells. Hind limb unloading in TNFΔARE mice significantly suppressed inflammation of the Achilles tendon compared with weight bearing controls. Erk1/2 signalling plays a crucial role in mechanotransduction-associated inflammation. Furthermore, new bone formation is strongly promoted at entheseal sites by biomechanical stress and correlates with the degree of inflammation.
Conclusions These findings provide a formal proof of the concept that mechanical strain drives both entheseal inflammation and new bone formation in SpA.
- Spondyloarthritis
- Inflammation
- Fibroblasts
Statistics from Altmetric.com
Introduction
Spondyloarthritides (SpA) are a group of chronic inflammatory disorders characterised by asymmetrical peripheral arthritis predominantly of lower limbs, and axial inflammation (sacroiliitis and spondylitis). The disease is typically accompanied by a variety of extra-articular manifestations, such as intestinal and ocular inflammation. In European countries, the overall incidence is estimated at 0.5%–2% of the Caucasian population, with onset frequently in the early adulthood.1 ,2 Inflammation of attachment sites of ligaments and tendons to bones, enthesitis, is a hallmark of SpA which distinguishes it from other inflammatory rheumatic disorders.3 In addition, SpA is also characterised by new bone formation evolving into ankylosis, or into the formation of enthesophytes that also appear to originate from these insertion sites.4 Radiographic progression of disease reflecting structural damage is characterised by new bone formation leading to sacroiliac and spinal ankylosis. Both inflammation and progressive structural damage contribute to the burden of disease.5
The entheses are subjected to repetitive biomechanical stressing forces that are applied during the course of normal muscle, ligament and tendon action; this suggests a link between biomechanical stress and SpA. McGonagle et al6 proposed an enthesitis-based model for the pathogenesis of SpA where interactions between biomechanical factors and the innate immune response (eg, to bacterial products) may lead to disease. Nevertheless, the enthesitis concept has remained controversial as synovitis and bone marrow inflammation are also typical signs of active SpA. The anatomical proximity and the ample molecular and cellular communications between these tissues suggest their functional relationship in the pathogenesis of synovitis in SpA with the disease processes defined in the context of the synovio–entheseal complex.7
Within this microenvironment, it has become clear that in SpA patients, new bone formation often occurs in close relationship with the entheses. Spinal syndesmophytes develop along the anterior intervertebral ligaments, and bony spurs are formed at the Achilles tendon and plantar fascia.
Interestingly, the process of inflammation and subsequent bone erosion appeared in anatomically distinct sites as compared with new bone formation at the Achilles’ enthesis.8 Erosions preferentially developed in regions undergoing compression, whereas spur formation occurred in regions prone to tensile forces. These anatomical data further corroborate current paradigms that strongly suggest a molecular uncoupling of inflammation and new bone formation in SpA. Despite a proven efficacy on inflammatory signs and symptoms, treatment with tumour necrosis factor (TNF) blocking agents in mouse models and in patients does not substantially affect progression of ankylosis.9–11
Currently, it remains elusive whether inflammation and new bone formation are closely linked or rather uncoupled, and why these processes colocalise at entheseal sites.
In the current study, we hypothesised that biomechanical factors drive inflammation and new bone formation at entheseal sites. This hypothesis was studied in the TNFΔARE mouse model in which chronic and deregulated TNF production leads to arthritis and a Crohn's-like ileitis.12–14 We previously argued that this model, depending on endogenous TNF, involving peripheral and axial joints and presenting extra-articular manifestations such as bowel inflammation, is a true translational model of SpA.13 Here, we observed that enthesitis is an early feature of this murine model, and that its development is dependent upon biomechanical strain. Bone new formation at entheseal sites, another prominent feature of SpA, was also associated with weight bearing and biomechanical strain. Altogether, these data substantiate the hypothesis that entheseal inflammation and new bone formation are driven by biomechanical strain.
Materials and methods
Animals and housing
TNFΔARE mice were provided by Kollias G.12 CAIA was induced with ArthritoMab from MD Biosciences according to the manufacturer's instructions. All mice were bred and housed in specific pathogen-free conditions in accordance with the general recommendations for animal breeding and housing, and all experiments were conducted in conditions according to the Ethical Committee of Animal Welfare. For treatment with MAPK inhibitors, mice were injected daily with 50 μg/g of PD98059, a selective Erk1/2/MEK1 inhibitor to inhibit Erk1/2 signalling, or SB203580 for p38 blockade (both LC Laboratories) intraperitoneally, or with DMSO as a negative control.
Histology
Murine paws and spine were dissected, fixed in 4% formaldehyde, decalcified in 0.5 M ethylenediamine tetraacetic acid (Sigma-Aldrich, St Louis, Missouri, USA) at 4° or in 5% formic acid until bones were pliable. Paraffin sections were stained with H&E for evaluation of inflammation and bone erosions, or with safranin-O for cartilage appreciation. Enthesitis of Achilles tendon was evaluated by two blinded assessors, and scored on three parameters, infiltrate in the Achilles tendon, calcaneal erosion and exudate at the synovio–entheseal complex, each ranging from 0 (normal) to 3. A composite score was built from these parameters. In addition, digital image analysis was performed by AxioVision software (Carl Zeiss MicroImaging GmbH, Göttingen, Germany). Extent of inflammation within the synovio–entheseal complex was normalised to the size of the growth plate. In a subgroup of mice from tail suspension experiments, inflammation in front paws was also evaluated as follows: absence (0) or presence (1) of dactylitis and enthesitis.
Western blotting
Achilles tendons were dissected on ice, snap frozen and lysed in laemli buffer by a bead-beating procedure to liberate proteins. Immunoblotting using chemiluminescent detection for phospho-Erk1/2 and total Erk1/2 was conducted as previously described. Briefly, equal amounts were loaded on precast 10% SDS-PAGE Criterion gels (Bio-Rad Laboratories, Hercules, California, USA). Equal loading was verified by Ponceau S staining (data not shown). MagicMark (Invitrogen, Paisley, UK) protein standards were run as Mw markers. Strips were focused for 35 kVh and SDS-PAGE was performed at 200 V for approximately 1 h. Proteins were transferred to nitrocellulose membranes (Bio-Rad Laboratories). The resulting membranes were immunoblotted with antiphospho-Erk1/2 or antitotal Erk1/2 (Cell Signaling Technology, Danvers, Massachusetts, USA), followed by antirabbit horseradish peroxidase-conjugated Ab and enhanced chemiluminescence (Pierce, Rockford, Illinois, USA). Chemiluminescence images were recorded using the VersaDoc-imaging system (Bio-Rad Laboratories). Densitometric analysis of the images was performed by Quantity One Software v 4.4.0 (Bio-Rad Laboratories).
Magnetic resonance imaging
Wild type and littermate TNFΔARE mice were anaesthetised with isoflurane (IsoFlo, Abbott, Illinois, USA) administered in a mixture of 30% O2 and 70% N2. During immobilisation, isoflurane levels were kept at 3%. The dose was gradually lowered to 1.5%. Mice were submitted to a longitudinal MRI study at age of 1, 2, 3 and 5 months. MRI was performed on a 9.4 Tesla MR system (BRUKER, Ettlingen, Germany). Coronal T1-weighted 2D images of the sacroiliac joints were acquired with a FLASH sequence: TE=3.4 ms, TR=200 ms, FA=40°, FOV=15 mm, image matrix (256×256), 16 slices, slice thickness =0.5 mm. T2-weighted images were acquired with a RARE sequence: TE=36 ms, TR=3000 ms, ETL=8, FOV=19.2 mm, image matrix (256×192), 8 slices, slice thickness =1 mm. To compare the T2-weighted signal intensities of the iliac bone among animals and age, relative signal intensities of the iliac bone with respect to the mean signal intensity of the muscle tissue close to the ilium bone were calculated.
Micro-CT acquisitions
Samples (n=36) were scanned in a GE Healthcare eXplore Locus SP preclinical micro-CT specimen scanner (GE Medical Systems, London, Ontario) with the following acquisition parameters: 80 kV tube voltage, 80 microamperes tube current, 1×1 detector binning, three times magnification, 500 projections acquired over 200°s using a 0.4° step size, 3000 ms exposure time and frame averaging of 4. The acquired projections were reconstructed into a 500×500×940 matrix, with 8-µm voxel size, by proprietary software (eXplore Reconstruction Utility, GE Medical Systems, London, Ontario) using a Feldkamp-type algorithm with Parker's weighting function.15 Each reconstructed micro-CT image was visually and quantitatively analysed by MicroView software (GE Medical Systems). Quantitative analysis was done by drawing a volume of interest. Bone mineral density was calculated inside this region after measuring the threshold for non-bone versus bone tissue, with the threshold value automatically determined.
A second set of micro-CT scans were performed at the Centre for X-ray Tomography of the Ghent University.16 To improve soft tissue contrast, the mice legs were immersed in a 4% HgCl2 aqueous solution for 24 h before scanning.31 The mice legs were scanned using the transmission head of a dual head x-ray tube from Feinfocus (FXE 160.51), operated at 120 kV, and a Varian 2520 V Paxscan flat panel detector. A beam filtration of 1 mm aluminium was applied. For each scan, 1441 projections were recorded, covering 360°, with an exposure time of 2 s per projection. The resulting voxel size was 6.5 µm. Reconstruction of the projection data was done using Octopus, a reconstruction software package developed at UGCT.17
Statistics
The non-parametric Kruskal–Wallis test or parametric ANOVA was used for comparison of three groups. The Mann–Whitney test was used for comparison of two groups. Results were considered statistically significant with two-sided p values <0.05. Regression analysis was performed to determine the correlation between osteophyte size and clinical score, irrespective of tail suspension, with R² the adjusted correlation coefficient.
Results
Features of spondyloarthritis in TNFΔARE mice
As the precise sites where inflammation originates in SpA remain a matter of debate, a detailed clinical and histological study was undertaken in TNFΔARE mice. Therefore, mice were scored every other day for arthritis development, and were sacrificed at different ages (4, 8, 12, 24 weeks) for histological assessment. In hind paws of TNFΔARE mice, inflammation initiated at approximately 4 weeks of age within the collateral ligaments of interphalangeal joints (figure 1B), within the synovio–entheseal complex of the Achilles tendon (figure 1E,F) and the greater trochanter ligament of the hip (figure 1H,I). By the age of 6–8 weeks, inflammation spreads out into the synovium with pannus formation, and finally involves the entire joint, with bone erosions (figure 1C). Thus, articular inflammation initiates within the entheseal regions in TNFΔARE mice.

Features of spondyloarthritis in TNFΔARE mice: entheseal inflammation. (A) Hind paw distal interphalangeal joint of healthy control mouse. (B) Early signs of enthesitis in hind paw of TNFΔARE mouse. (C) Advanced erosive stage. (D) Achilles tendon of healthy control mouse. (E and F) Achilles enthesitis in TNFΔARE mouse. (G) Hip joint of healthy control mouse. (H and I) Hip joint of TNFΔARE mouse, arrow head points at inflammation around the greater trochanter. (J) Sacroiliac joint of a healthy control mouse. (K) Sacroiliac joint of young TNFΔARE mouse. (L) Advanced stage sacroiliitis in TNFΔARE mouse, white arrows point at erosions. Staining with H&E. Original magnification×100, except D, E, G and H original magnification×40. Access the article online to view this figure in colour.
Another interesting hallmark of the disease is the development of spinal inflammation with sacroiliitis (figure 1J–L).13 The joints between the sacrum and the pelvic ilium bone are joined by several ligaments and consist of a lower fibrocartilagenous part and an upper ligamentous part. As such, this region is also prone to mechanical strain. As sacroiliac joints are difficult to assess clinically, we performed a longitudinal in vivo MRI study, in which TNFΔARE mice and controls were scanned monthly, starting at the age of 4 weeks, and each month both T1 and T2 sequences were run (figure 2A). On T1 sequences, progressive narrowing of joint space and irregular articular surfaces reflected progressive structural damage. On T2 sequences, the relative signal intensity of the iliac bone was calculated for each mouse (figure 2B). The signal intensity in control mice clearly decreased with age because of an increase in mineral content (63% decrease at 5 months compared with 1 month). The signal intensity in TNFΔARE mice only decreased by 29% between 1 and 5 months of age. This may reflect an increased water content (or bone marrow oedema) and a smaller mineral content in TNFΔARE mice. Alternatively, the differences in signal may also reflect the persistence of red marrow (ie, increased cellularity) seen in TNF overexpression models.18 The two-way analysis of variance (ANOVA) of relative signal intensity of the iliac bone values demonstrated both a significant main group effect (TNFΔARE mice vs controls) and age effect. At the age of 2, 3 and 5 months, the relative signal intensities of the TNFΔARE mice were significantly higher compared with the controls (figure 2B). Detailed images obtained by this in vivo imaging approach provide a valuable tool to study sacroiliitis development and allow the monitoring of therapeutic interventions in animal models for SpA. Collectively, these data highlight that inflammation occurring in TNFΔARE mice is highly reminiscent of human SpA.
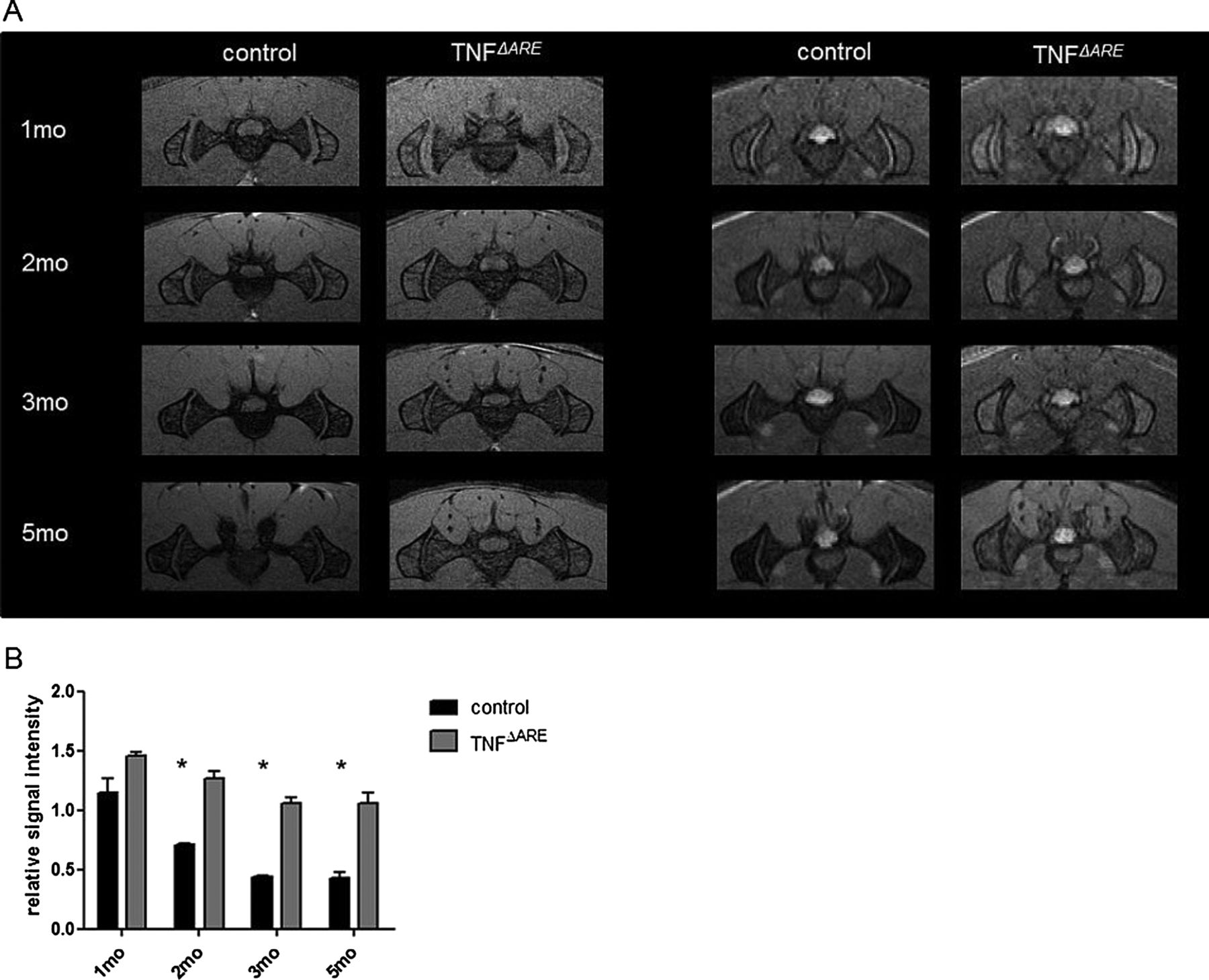
Consecutive monthly in vivo MRI of sacroiliac joints of healthy control and littermate TNFΔARE mice. (A) T1-weighted images (left panel) illustrate the development of sacroiliitis in TNFΔARE mice: joint space narrowing, irregular surfaces. T2-weighted images (right panel) illustrate the persistence of red marrow and general osteopenia in TNFΔARE mice. (B) Graph gives the relative T2-weighted signal intensities of the ilium with respect to a neighbouring muscle control region for control and TNFΔARE mice and for different ages. A two-way ANOVA demonstrates differences in relative signal intensities between the group of control mice and TNFΔARE mice, and for the different ages of the mice (*=p<0.05).
Enthesitis occurs independent of mature T and B cells
It was recently shown that interleukin (IL)-23 responsive CD3+CD4–CD8– entheseal resident T cells are essential to induce enthesitis in an IL-23 overexpression model.19 To assess the role of adaptive immunity, we backcrossed TNFΔARE mice on to RAG-1 deficient animals that have no mature T cells, and assessed severity of enthesitis. In RAG-1 deficient TNFΔARE mice, incidence of enthesitis was similar to that in T cell sufficient TNFΔARE mice and occurred in all mice examined (figure 3B,C). This finding highlights that both T cell dependent and independent mechanisms, most likely involving stromal cells, can contribute to enthesitis. Our data corroborate the previous role for stromal cells in TNF driven joint inflammation.13

Inflammation at the synovio–entheseal complex is driven by biomechanical factors in TNFΔARE mice. (A) Healthy control mouse: overview of ankle. (B) Achilles tendon from a RAG-1 deficient TNFΔARE mouse. (C) Achilles tendon from RAG-1 sufficient (heterozygous) littermate TNFΔARE mouse. (D) Achilles tendon of 4-week-old healthy control mouse. (E) Non-tail suspended TNFΔARE mouse. (F) Littermate TNFΔARE mouse subjected to tail suspension for 14 days continuously. (G) Composite histology score demonstrates reduced severity of enthesitis upon tail suspension, *=p<0.05. (H) Distal interphalangeal joint in front paw of a non-tail suspended TNFΔARE and (I) tail suspended TNFΔARE mouse. All original magnification ×40. (J) Graph demonstrates inflammation score in front paws from a subgroup of mice from tail suspension experiments, N=4 in each group, values are mean and SEM. Access the article online to view this figure in colour.
Hind limb unloading inhibits development of enthesitis in TNFΔARE mice
We hypothesised that continuous mechanical strain caused by normal animal activity could be at the origin of enthesitis in this model. The Achilles tendon was chosen as the focus of interest for this study. To address this we used an established model for hind limb unloading, tail suspension.20 ,21 In specially designed cages, TNFΔARE mice and healthy controls were tail suspended, and clinical signs of arthritis were assessed daily. Hind limb unloading was initiated before the known onset of illness, before 4 weeks of age, in a preventive setting. Following a 14-day period of unloading, none of the mice subjected to this procedure displayed clinical signs of peripheral arthritis of ankles and hind paws, whereas all control mice that were not tail suspended, did. Furthermore, by detailed histological analysis, we detected only minor inflammatory changes in tail suspended TNFΔARE mice, but significantly more inflammation in their weight bearing controls (figure 3D–G). In a subgroup of mice, arthritis was also evaluated in front paws. In general, severity of inflammation did not differ between weight carrying front paws of tail-suspended and control mice (figure 3H–J). These data strongly suggest that the onset of enthesitis in this animal model is driven by mechanical load on hind paws.
Mechanical stress triggers Erk1/2 signalling in TNFΔARE mice
Activation of mitogen-activated protein kinase (MAPK) signalling pathways was previously demonstrated as a response to stretch in a variety of stromal cell types.22–25 Therefore, we performed a tail suspension study, whereby TNFΔARE mice were subjected to tail suspension for 7 days, again in a preventive setting, before the onset of enthesitis. Tail suspended mice did not show signs of clinical arthritis, as compared with weight bearing controls. Seven days later, half of the group of mice were allowed to walk around for 10 min before they were sacrificed. Induction of phosphorylated-Erk1/2 (P-Erk1/2), phosphorylated-p38 (P-p38) and phosphorylated Jnk (P-Jnk) was assessed by western blotting on Achilles tendon lysates. As figure 4A indicates, in Achilles tendon lysates from wild type mice subjected to tail suspension, P-Erk1/2 was comparable with weight bearing wild type mice, while the reintroduction of mechanical strain during 10 min resulted in a small albeit not significant induction of P-Erk1/2. By contrast, in TNFΔARE Achilles tendon, a manifest increase of P-Erk1/2 could be detected upon 10 min of weight bearing, with significant induction of P-Erk2 on densitometry (figure 4B). No significant signal of P-p38 was detected, and P-Jnk was not detected at all (not shown).

Erk1/2 signalling is involved in enthesitis development in TNFΔARE mice. (A) Western blotting on Achilles tendon lysates of 4-week-old tail suspended (7 days) and non-tail suspended healthy control and TNFΔARE mice. Half of the group of tail suspended mice was allowed to walk around for 10 min before they were sacrificed. Figure shows one representative experiment out of three. (B) Densitometric analysis of the images was performed by Quantity One Software. N=5 mice per condition, *=p<0.05. (C) H&E staining of the Achilles tendon and ankle of 4-week-old TNFΔARE mice treated with PD98059, SB203580 or DMSO for 7 days. (D) Inflammation within the synovio–entheseal complex (SEC) was scored by digital image analysis, and is shown as the ratio of the inflamed area within the SEC and the area of the growth plate. (E) The extent of erosion of the calcaneal bone was scored by two blinded assessors. N=3–5 mice in each group. Values are means and SEM. *=p<0.05. Access the article online to view this figure in colour.
MAPK inhibition interferes with tendinitis development
Small molecular inhibitors of MAPK pathways whose activity has previously been assessed in vitro and in vivo were selected. The selective Erk1/2/MEK1 inhibitor PD98059 was used to inhibit Erk1/2 signalling, and SB203580 was used for p38 pathway inhibition26 ,27 (and own unpublished data); 4-week-old TNFΔARE mice were injected intraperitoneally with inhibitors (50 µg/g) or vehicle (dimethylsulphoxide (DMSO)) for seven consecutive days, and development of arthritis was assessed daily. Detailed histological analysis of H&E stained ankles using digital image analysis showed marked reduction in inflammation within the synovio–entheseal complex in mice treated with PD98059 (figure 4C,D). In addition, there was significantly less calcaneal bone erosion in mice treated with PD98059 versus control mice (figure 4E). These findings indicate that blockade of the Erk1/2 signalling pathway altered the development of enthesitis. Inflammation within the ileum, another disease manifestation in TNFΔARE mice, was not altered significantly upon treatment with MAPK inhibitors (data not shown).
New bone formation
New bone formation, an important feature of human spondyloarthritis, does not appear in TNFΔARE mice, consistent with previous work by Diarra et al28 performed in the human TNF transgenic (hTNFtg) mouse model. TNF was shown to be the major inducer of Dickkopf-1 (DKK-1), a regulatory molecule of the Wnt pathway, which operated as a brake on new bone formation. In addition, blockade of DKK-1 resulted in osteophyte formation within arthritic joints of hTNFtg mice. Therefore, we focused on the collagen-antibody-induced arthritis model (CAIA) to study new bone formation in relation to mechanical stress. CAIA was induced in susceptible DBA/1 mice as described.29 ,30 As early as 7 days after immunisation, CAIA mice exhibit enthesitis which eventually evolves into a destructive polyarthritis19 (data not shown). Osteophyte formation occurs after resolution of the inflammatory phase. When a full blown clinical arthritis was established at day 10 after immunisation, half of the group of mice were tail suspended for 28 days to prevent mechanical loading on hind paws, and half were kept in normal cages. In tail suspended mice, osteophytes were significantly smaller compared with control mice, as shown by histology and micro-CT imaging (figure 5A,B). In general, osteophyte size correlated with the severity of inflammation. Mice with only minimal clinical arthritis developed no osteophytes when tail suspended, whereas mice with higher arthritis scores developed significantly smaller osteophytes when tail suspended compared with non-tail suspended mice (figure 5C,D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
New bone formation is driven by mechanical strain. (A and B) Micro-CT images and corresponding H&E staining of the region within the square from healthy control, non-tail suspended and tail suspended collagen-antibody-induced arthritis mice. (C) Osteophyte size was scored by two blinded assessors. Graph shows osteophyte size for mice with minor (0–1) or major (2–3) clinical arthritis score (one paw). N=10 mice in each group, *=p<0.05. (D) Correlation of osteophyte size with clinical score. R2=0.28, p=0.03. Access the article online to view this figure in colour.
New bone formation occurred at distance from the joint articular surfaces. Moreover, immersion of the entire paw in mercury chloride (HgCl2), which allows uptake in soft tissues such as tendons,31 followed by high resolution CT, revealed that these osteophytes are formed mainly at entheseal sites (see online supplementary figure S1 and S2). Collectively, these findings underscore a role for mechanical strain in new bone formation at entheseal sites.
Discussion
It has been a great matter of debate where disease originates in SpA patients since, during the course of disease, synovium, enthesis as well as bone marrow can become involved. Due to their location at the interface of ligaments and bone, entheses are especially prone to mechanical forces. It is generally accepted although not formally proven that both mechanical and inflammatory factors can trigger enthesitis, with the enthesitis-based origin of SpA as a formal hypothesis.6 According to an occupational study by Ward et al,32 physical activities that required repetitive strain such as stretching, bending or twisting, as well as exposure to whole body vibration, were associated with more radiographic damage in ankylosing spondylitis. Yet, experimental evidence supporting these theories was lacking. We provide evidence for a mechanotransduction-associated origin of enthesitis and new bone formation at entheseal sites.
First, enthesitis is an early disease feature in TNFΔARE mice. Hind limb unloading prevented the onset of Achilles enthesitis in TNFΔARE mice. Front paws of tail suspended TNFΔARE mice showed similar arthritis severity compared with weight bearing controls, although the body weight of these mice was loaded on the front paws. However, careful observation revealed that tail suspended mice tended to move less than the control group, which provides an explanation for the contra-intuitive finding that severity of inflammation in front paws does not increase after tail suspension and hind limb unloading. We hypothesise that minimal biomechanical stress is sufficient to induce a sustained inflammatory response. Thus, biomechanical stress rather functions as an on/off switch and there is not necessarily a linear relationship between the amount of stress and the degree of inflammation, at least not above a given level of biomechanical stress. The resulting minor increase, if any, may not be detected by the scoring system, which has limited power.
Therefore, mechanical strain may be considered the trigger of an inflammatory process, and provides an explanation for the unsolved paradigm of enthesitis development in SpA.
It was previously demonstrated that in the presence of chronic TNF overexposure, signalling through TNFRI in synovial fibroblasts and intestinal myofibroblasts appears to be sufficient to develop arthritis and Crohn’s-like ileitis in TNFΔARE mice.13 Therefore, crosstalk between the myeloid cell compartment and stromal cells such as fibroblasts is critically involved in the development of both the articular and the intestinal phenotype. Importantly, mature T cells are not involved in enthesitis development in TNFΔARE mice, as demonstrated in RAG-1 deficient mice. This highlights that several pathways exist to induce enthesitis, some of which rely on IL-23R+ innate-like entheseal resident cells,19 while others are strictly dependent upon stromal cell function. In more complex situations such as in SpA patients, both T cell dependent and independent mechanisms may exist and could have additive effects.
Mechanical stress is sensed by cells via mechanoreceptors such as integrins, and transmitted through MAPK signalling pathways.33 In this study, activation of Erk1/2 signalling pathways in Achilles tendon lysates could be efficiently prohibited by hind limb unloading, while reintroduction of mechanical strain resulted in immediate phosphorylation of Erk1/2 (within 10 min), both in wild type and TNFΔARE tendons. Small molecular MAPK inhibitors were proven effective in animal models for arthritis, and may be promising therapeutic targets in treatment of inflammatory diseases.34 ,35 However, limited data are available on treatment options of enthesitis in particular. In the present study, the effects of an Erk1/2 inhibitor and a p38 inhibitor on enthesitis development were evaluated. With Erk1/2 inhibition, we detected a clear decrease in inflammation within the synovio–entheseal complex; in addition, there was a significant reduction in calcaneal bone erosion.
The MAPK pathways have previously been shown to modulate TNF biosynthesis, and TNF signalling is involved both upstream and downstream of MAPK activation.36 In particular, the adenosine uracil (AU) rich elements within the TNF transcript are known targets for p38-mediated activation of TNF translation upon lipopolysaccharide stimulation.12 In addition, the Erk1/2 pathway requires the presence of ARE for post-transcriptional regulation, that is, nuclear export of the TNF message.37 In wild type mice, MAPK blockade in general results in a dose dependent inhibition of TNF translation. As these targets are deleted in TNFΔARE mice, MAPK blockade would be unable to inhibit TNF translation via this pathway. Therefore, the observed biological effects of Erk1/2 blockade in TNFΔARE mice may even be underestimated and partly TNF independent. However, data from other models suggest that inhibition of MAPK has complex effects. In the ankylosing enthesitis model in DBA/1 mice, inhibition of p38 accelerated ankylosis rather than inhibiting arthritis, although in vitro data suggested an inhibitory effect of the compound used on transforming growth factor and bone morphogenetic protein induced chondro- and osteogenesis.38
A second hallmark of spondyloarthritis is the occurrence of new bone formation leading to ankylosis. However, in TNFΔARE mice, new bone formation could never be demonstrated, presumably due to continuous and deregulated TNF production that functions as a brake on this process through for example DKK1, which is a potent inhibitor of Wnt signalling. In addition, new bone formation is much less pronounced in the CB57BL/6 background than on other backgrounds such as DBA/1. Successful inhibition of inflammation in mouse models and in patients, for instance with TNF blocking agents, does not affect ankylosis or radiographic progression.9–11 However, recent data suggest that long term treatment may be effective on structural changes.39 ,40 The sequence of inflammation followed by new bone formation is not formally proven, although there may be a preference for syndesmophytes to develop at inflamed vertebral edges in human SpA.41 ,42 In addition, recent MRI studies suggested that new bone formation more likely develops in advanced inflammatory spinal lesions. In these lesions, resolution of inflammation results in fat metaplasia and ultimately in new bone formation.43 ,44 Here, we demonstrated that in CAIA mice, osteophytes are significantly smaller when weight bearing is prohibited, supporting a role for mechanical strain in their development. In addition, tail suspension was able to prevent the development of osteophytes when only minimal signs of inflammation were evident, and could minimise osteophyte size in more advanced arthritis. In general, osteophyte size correlated with the severity of inflammation. Although this particular aspect was not the focus of this study, it indirectly supports the relationship between inflammation and new bone formation.
In summary, these findings comprise the first formal proof of a new paradigm that mechanical forces are the underlying cause of enthesitis and new bone formation in SpA. This opens avenues for a new field of mechanotransduction-associated pathways in SpA.
Acknowledgments
The authors wish to thank C Van Hove at INFINITY for performing and analysing micro-CT images and L Van Praet for statistical analysis.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
- Data supplement 2 - Online video
Footnotes
Handling editor Tore K Kvien
-
Contributors All authors contributed to the conception and design of the study, acquisition, analysis or interpretation of data, and drafting or revising the article critically for intellectual content. All authors approved the final version for publication.
-
Funding This work is supported by the Fund for Scientific Research—Flanders, by a concerted action grant from Ghent University, and by the Special Research Fund of the Ghent University (BOF, GOA 01G01008). DE is part of a multidisciplinary research platform (MRP) of Ghent University and is supported by an Interuniversity Attraction Pole (Project P7/07).
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Data sharing statement Data not shown are available upon email request to the corresponding author.