Article Text

Abstract
Objectives: To investigate the role of reactive oxygen species (ROS) in the development of the various patterns of systemic sclerosis (SSc) and the mechanisms of ROS production by endothelial cells and fibroblasts.
Methods: Production of hydrogen peroxide (H2O2), nitric oxide (NO) and cellular proliferation were determined following incubation of endothelial cells and fibroblasts with 56 SSc and 30 healthy sera. Correlations were established between those markers, the type and the severity of the clinical involvements, and the response to treatment. The factors leading to ROS production were determined.
Results: H2O2 production by endothelial cells and fibroblasts was higher after incubation with SSc sera than with normal sera (p<0.001) and with sera from SSc patients with severe complications than sera from other patients (p<0.05). Sera from patients with lung fibrosis triggered the proliferation of fibroblasts more than other SSc sera (p<0.001), whereas sera from patients with vascular complications exerted no proliferative effect on fibroblasts, but inhibited endothelial cell growth (p<0.05) and induced NO overproduction (p<0.05). Bosentan reduced NO release by 32%, whereas N-acetylcystein potentiated 5-fluorouracil (5FU) to inhibit fibroblast proliferation by 78%. Those serum-mediated effects did not involve antibodies but advanced oxidation protein products that selectively triggered cells to produce H2O2 or NO.
Conclusions: SSc sera induce the production of different types of ROS that selectively activate endothelial cells or fibroblasts, leading to vascular or fibrotic complications. Assaying serum-induced ROS production allows clinical activity of the disease to be followed and appropriate treatments to be selected.
- AOPPs, advanced oxidation protein products
- DAF2-DA, 4,5-diaminofluorescein diacetate
- 5FU, 5-fluorouracil
- H2-DCFDA, 2′,7′-dichlorodihydrofluorescein diacetate
- HCs, healthy controls
- HUVECs, human umbilical venous endothelial cells
- NAC, N-acetyl-l-cysteine
- NO, nitric oxide
- PAH, pulmonary arterial hypertension
- ROS, reactive oxygen species
- RS, restrictive syndrome
- SSc, systemic sclerosis
- VAs, vascular abnormalities
- systemic sclerosis
- advanced oxidation protein products
- reactive oxygen species
- endothelial cells
- fibroblasts
Statistics from Altmetric.com
- AOPPs, advanced oxidation protein products
- DAF2-DA, 4,5-diaminofluorescein diacetate
- 5FU, 5-fluorouracil
- H2-DCFDA, 2′,7′-dichlorodihydrofluorescein diacetate
- HCs, healthy controls
- HUVECs, human umbilical venous endothelial cells
- NAC, N-acetyl-l-cysteine
- NO, nitric oxide
- PAH, pulmonary arterial hypertension
- ROS, reactive oxygen species
- RS, restrictive syndrome
- SSc, systemic sclerosis
- VAs, vascular abnormalities
- systemic sclerosis
- advanced oxidation protein products
- reactive oxygen species
- endothelial cells
- fibroblasts
Systemic sclerosis (SSc) is a connective tissue disorder of unknown aetiology; the main features are vascular hyper-reactivity and fibrosis of skin and visceral organs.1,2
SSc is characterized by a large variety of clinical patterns. Although genetic and environmental factors are known to play a role in the initiation and progression of the disease, the mechanisms that determine the clinical manifestations remain unclear.3–5
Recent reports have suggested that oxidative stress could be involved in the pathogenesis of SSc,6–9 and spontaneous production of reactive oxygen species (ROS) by skin fibroblasts from SSc patients has been observed.6,10
Our aim was to determine whether serum factors can mediate an oxidative stress that activates endothelial cells and fibroblasts in SSc patients. The involved ROS and their respective effects on both types of cell have been characterized in order to determine the mechanisms that lead to a vascular or a fibrotic form of the disease. In addition, various drugs and antioxidant molecules have been tested for their counteracting effects on the oxidative stress.
PATIENTS AND METHODS
Patients and sera
Fifty-six SSc patients were enrolled in the study. The clinical and serological characteristics of these SSc patients are summarized in table 1.
Clinical and serological characteristics of the 56 SSc patients
The onset of the disease was defined as the time of the first non-Raynaud’s phenomenon. To be eligible for the study, patients had to fulfill the American College of Rheumatology criteria and/or the Leroy & Medsger criteria for SSc.11–13 Limited cutaneous SSc was defined by skin thickening in areas solely distal to the elbows and knees, and diffuse cutaneous SSc was defined by the presence of skin thickening that was proximal, as well as distal, to the elbows and knees.12
Interstitial lung disease was assessed by chest high-resolution computed tomodensitometry and pulmonary function test values. A restrictive syndrome (RS) was defined by a reduction (<75%), compared with values obtained from a well-defined population of normal subjects, matched for gender, age, height and ethnic origin using similar test protocols in both forced expired volume in one second (FEV1) and forced vital capacity (FVC), and a FEV1/FVC ratio >75%.
Vascular abnormalities (VAs) were defined as digital ulcers or scars of ulcers related to digital ischaemia or pulmonary arterial hypertension (PAH) defined by systolic pulmonary arterial pressure >40 mmHg following echocardiography and/or mean pulmonary arterial pressure >25 mmHg following right-heart catheterization in the absence of RS. Twelve patients presenting with active digital ulcers, and/or scars of digital ulcers, and six patients with PAH in the absence of RS, were classified as patients with VAs. One patient had both digital ischaemia and PAH, and this was taken into account in both groups. The eight patients with PAH associated with a RS were not included in the group of patients with vascular primitive PAH in order to form a homogenous group of patients with primitive vascular abnormalities. Only 1 of 56 patients had both RS and VAs, and this was taken into account in both groups.
As no sample contemporary of a renal crisis was available, this complication was not used to classify patients in this study. Two patients had tendon friction rubs. None of the 56 patients had another connective tissue disease or cancer.
Normal sera were collected from 30 healthy adults (22 females and 8 males, mean age 44±17 years). All patients and controls gave their written informed consent.
Cells and chemicals
Human umbilical venous endothelial cells (HUVECs) were obtained by digestion of umbilical cords with 0.1% collagenase. NIH 3T3 fibroblasts were obtained from the American Type Culture Collection (Manassas, VA). HEp-2 cells were obtained from EuroBio (Les Ulis, France). Primary normal human dermal fibroblasts were a gift from A. Munnich (INSERM 781, Necker Hospital, Paris, France). All cells were cultured as previously reported.14,15
Measurement of hydrogen peroxide and nitric oxide production
Cells (8 × 103 per well) were seeded in 96-well plates (Costar, Corning, Inc, NY) and incubated with their respective growth medium alone for 12 hours at 37°C in 5% CO2. Culture media were then removed and cells pre-incubated with 50 μl 2′,7′-dichlorodihydrofluorescein diacetate (H2-DCFDA) diluted 1:1000 in phosphate-buffered saline (PBS) to test cellular hydrogen peroxide (H2O2) production or with 50 μl 4,5-diaminofluorescein diacetate (DAF2-DA) diluted 1:375 in PBS to test nitric oxide (NO) release. After 30 minutes, 50 μl of SSc or control sera were added, and H2O2 and NO productions were assayed spectrofluorimetrically after 6 hours (Fusion, PerkinElmer, Wellesley, MA, USA). Results were expressed in arbitrary units (AUs) per minute and per million cells.
In vitro cell proliferation assays
HUVECs, as well as NIH 3T3, human primary fibroblasts or HEp-2 cells (4 × 103 per well), were seeded in 96-well plates (Costar) and incubated with 50 μl of SSc or control serum and 150 μl of culture medium without fetal bovine serum (FBS) at 37°C in 5% CO2 for 48 hours. Cell proliferation was determined by pulsing the cells with [3H]thymidine (1 μCi/well) during the last 16 hours of culture. Results were expressed as absolute numbers of counts per minute.
Assay of serum anti-endothelial and anti-fibroblast antibodies
NIH 3T3 fibroblasts and HUVECs (4 × 104 cells per well) were incubated with 1:5 dilution of SSc or control sera for 1 hour at 4°C, then washed in PBS and incubated with 1:200 FITC-rabbit polyclonal anti-human IgG, A, M antibody (Dako, Glostrup, Denmark) for 1 hour at 4°C. Fluorescence intensity was determined spectrofluorometrically and expressed as AUs.
Assay of advanced oxidation protein products in sera
Advanced oxidation protein products (AOPPs) were measured by spectrophotometry as previously described.16 The assay was calibrated using chloramine-T. The absorbance was read at 340 nm on a microplate reader (Fusion, PerkinElmer, Wellesley, MA, USA). AOPP concentrations were expressed as μmol/l of chloramine-T equivalents.
ROS production by cellular AOPP
HUVEC protein extracts were obtained by incubation of HUVECs with 1% NP40 and protease inhibitors in PBS buffer. Proteins were oxidized by 1 mM HOCl for 1 hour or 1 mM peroxynitrites for 18 hours at room temperature, then dialysed overnight against PBS and tested for AOPP content. Endothelial cells and NIH 3T3 cells (8 × 103 per well) were incubated with 0.5 mg per well of either unoxidized or oxidized HUVEC extracts. Production of H2O2 and NO was assessed spectrofluorometrically using H2-DCFDA and DAF2-DA as described above.
Effects of drugs on serum-induced ROS production
SSc sera were incubated with either 50 μM bosentan (Actelion, Allschwil, Switzerland), 10 μM nifedipine (Bayer Pharma, Leverkusen, Germany), 50 μM d-penicillamine (Dexo, Saint Cloud, France) or 10 μg etanercept (Wyeth, Madison, USA) in PBS for 90 minutes at 37°C. PBS alone was used as control. HUVECs (8 × 103 per well), seeded in 96-well plates, were incubated with 50 μl of H2-DCFDA or DAF2-DA. After 30 minutes, the sera with drugs were added, and H2O2 and NO productions were assessed as above. In other experiments, HUVECs and NIH 3T3 cells (8 × 103 per well) were incubated with 1600 μM N-acetyl-l-cysteine (NAC), 400 μM reduced glutathione (GSH) or 10 U PEG-catalase for 18 hours at 37°C in 5% CO2. Medium and chemicals were removed and replaced by 50 μl of H2-DCFDA (for HUVECs and NIH 3T3 cells) or DAF2-DA (for HUVECs). After 30 minutes, 50 μl serum diluted 1:2 were added and ROS production was assessed as above. Results were expressed as percentages of ROS production versus untreated cells (100%).
Effects of drugs on serum-induced HUVECs and fibroblast proliferation
NIH 3T3 cells or HUVECs (4 × 103 per well) were incubated with SSc or control serum (1:8 v:v) in culture medium without FBS at 37°C in 5% CO2 for 48 hours. For NIH 3T3 cells, one of the following molecules was added: 1600 μM NAC, 400 μM reduced GSH, 10 μM nifedipine, 50 μM d-penicillamine, 50 μM bosentan, 10 μg etanercept, 10 U PEG-catalase or 25 μM 5FU (Dakota Pharm, Le Plessis Robinson, France), with or without 1600 μM NAC. For HUVECs, 600 μM NAC, 400 μM reduced GSH or 10 U PEG-catalase were added. Cell proliferation was assessed as described above. Results were expressed as percentages of cellular proliferation versus untreated cells (100%).
Longitudinal study of in vitro ROS production by HUVECs and of NIH 3T3 fibroblast proliferation
A longitudinal study was performed with the sera from 5 patients. Productions of H2O2 and NO, cell proliferation and serum AOPP concentration were measured as previously described.
Statistical analysis
Data were compared with the Student’s t unpaired, paired tests and Fisher’s exact test. When analysis included >2 groups, one-way ANOVA was used. Pearson’s r correlation coefficient was used to analyse the relationships between quantitative variables. A p value <0.05 was considered to be significant. Quantitative values higher than 2 SD above the mean obtained with control sera were considered to be positive.
RESULTS
Effects of SSc sera on ROS production and proliferation of fibroblasts, HUVECs and HEp-2 cells
HUVECs, NIH 3T3 fibroblasts or HEp-2 cells exposed to SSc sera released higher amounts of H2O2 than when exposed to normal sera (p<0.001 in all cases; fig 1A). In contrast, mean NO release was similar whether HUVECs, NIH 3T3 fibroblasts or HEp-2 cells had been incubated with sera from SSc patients or with normal sera (fig 1B).

Effects of 56 sera from SSc patients (solid symbols) and 30 sera from healthy controls (HCs) (open symbols) on ROS generation and cell proliferation. (A) Production of hydrogen peroxide (H2O2) and (B) Production of nitric oxide (NO) by human umbilical venous endothelial cells (HUVECs), NIH 3T3 fibroblasts and HEp-2 cells. (C) Cell proliferation determined by [3H]thymidine incorporation. (D) Primary normal human fibroblast proliferation. Error bars represent SEM. *Denotes statistical significance.
As ROS can modulate cell proliferation, we tested the effects of SSc sera on the proliferation of NIH 3T3 cells and human fibroblasts and HUVECs, and HEp-2 cells as control. SSc sera exerted a higher proliferative effect on NIH 3T3 fibroblasts (fig 1C) and on human fibroblasts (fig 1D) than normal sera (p = 0.03 and p = 0.001, respectively) and exerted an anti-proliferative effect on HUVECs when compared with normal sera (p<0.001). No difference between SSc sera and control sera was observed on HEp-2 cells (fig 1C). A strong correlation was observed between the proliferative effect exerted by SSc sera on normal human fibroblasts and on NIH 3T3 cells (r = 0.85; p<0.0001).
H2O2 production by NIH 3T3 fibroblasts was correlated with SSc serum-induced proliferation (r = 0.31, p = 0.02). The anti-proliferative effect observed on HUVECs was also correlated with the amounts of H2O2 produced by these cells (r = –0.30, p = 0.03).
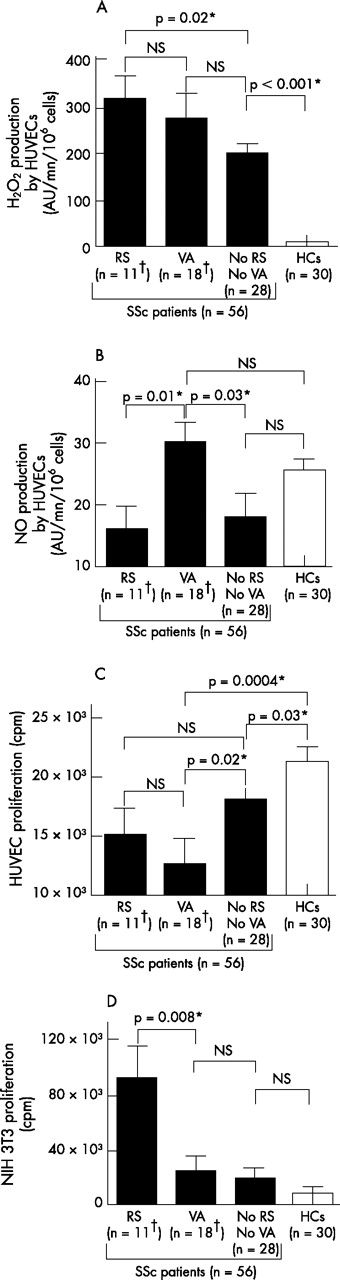
Correlations between clinical involvement of the disease and serum-induced ROS production and cellular proliferation
Sera from the 11 patients with a RS induced higher levels of H2O2 than sera from the 28 SSc patients with no RS and no vascular abnormality and from healthy subjects (p = 0.02 and p<0.0001; fig 2A). Conversely, sera from these 11 patients did not induce higher release of NO compared with sera from the two other groups (fig 2B). These 11 sera exerted no higher proliferative effect on HUVECs than sera from patients with digital ischaemia or vascular PAH, or from SSc patients with no RS and no vascular abnormality (fig 2C and data not shown). In contrast, these 11 sera exerted a significantly higher proliferative effect on NIH 3T3 fibroblasts compared with sera from patients with no RS and no vascular abnormality, from patients with digital ischaemia, with vascular PAH and from healthy subjects (p = 0.04; p = 0.04; p = 0.02; p = 0.008; fig 2D and data not shown).
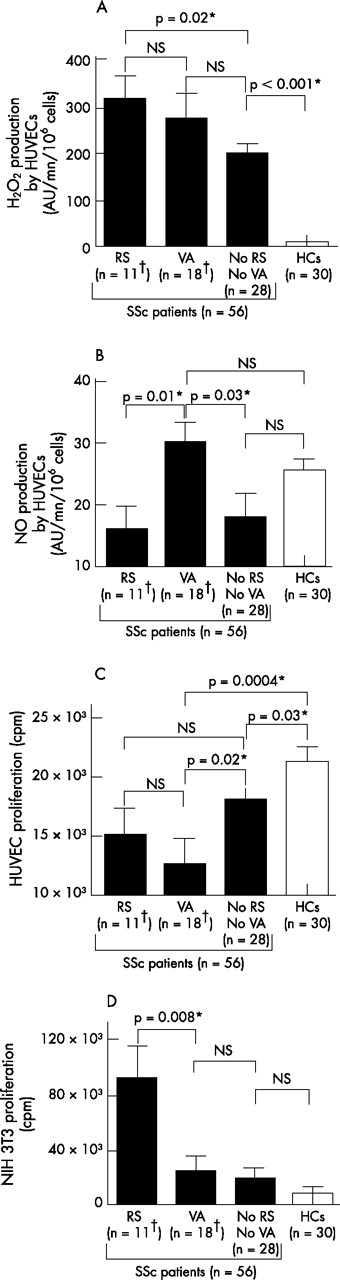
ROS production and cell proliferation induced by sera from SSc patients according to their clinical pattern. A total of 11 patients had a restrictive syndrome (RS); 18 had vascular abnormalities (VAs); 28 had none of these complications; 30 sera were collected from healthy controls (HCs). (A) Serum-induced production of hydrogen peroxide (H2O2) by human umbilical venous endothelial cells (HUVECs). (B) Serum-induced production of nitric oxide (NO) production by HUVECs. (C) Serum-induced proliferation of HUVECs. (D) Serum-induced proliferation of NIH 3T3 fibroblasts. Error bars represent SEM. *Denotes statistical significance. †One patient had both a RS and VAs.
Sera from the 18 patients with vascular abnormality (11 patients with digital ischaemia, 5 with PAH and 1 with both involvements) induced a higher release of NO by HUVECs than sera from the 11 patients with a RS and from the 28 SSc patients with no RS and no vascular abnormality (p = 0.01 and p = 0.03, respectively; fig 2B). These 18 sera exerted no effect on the growth of NIH 3T3 fibroblasts when compared with sera from the 28 SSc patients with no RS and no vascular abnormality and with sera from healthy controls (fig 2D), but abrogated the proliferation of HUVECs more strongly than the sera from these two other groups of individuals (p = 0.02 and p = 0.0004, respectively; fig 2C). Among these 18 patients with vascular abnormality, no difference was observed between patients with digital ischaemia and those with PAH either in H2O2 and NO induction or in modulation of cell proliferation. Nevertheless, the sera from the 12 patients with digital ischaemia abrogated the proliferation of HUVECs more strongly than the sera from the 28 SSc patients with no RS and no vascular abnormality (p = 0.02 and p = 0.18, respectively; data not shown). This significant difference was not observed with sera from the six patients with PAH.
Conversely, 50% of the 12 patients with significant serum-induced fibroblast proliferation versus controls presented with a RS that was detected in only 11.4% of the other SSc patients (p = 0.007). Fifty-five percent of patients with significant serum-induced anti-proliferative effect on HUVECs suffered from digital ischaemia that was present in only 13% of the other SSc patients (p = 0.007).
No difference was observed in NO and H2O2 release and on cell proliferation between patients with diffuse or limited cutaneous SSc.
Serum AOPPs are responsible for ROS production and modulate fibroblast proliferation
We then characterized the serum factors responsible for the oxidative stress. We observed no correlations between anti-centromere, anti-topoisomerase 1 (table 1), anti-endothelial cell (24% SSc sera) or anti-fibroblast antibodies (28% SSc sera), and H2O2 or NO production by endothelial cells or fibroblasts (p⩾0.13 in each case). In addition, the depletion of serum IgG by protein G (assessed by nephelometric assay) affected neither serum-induced ROS production nor fibroblast proliferation (data not shown).
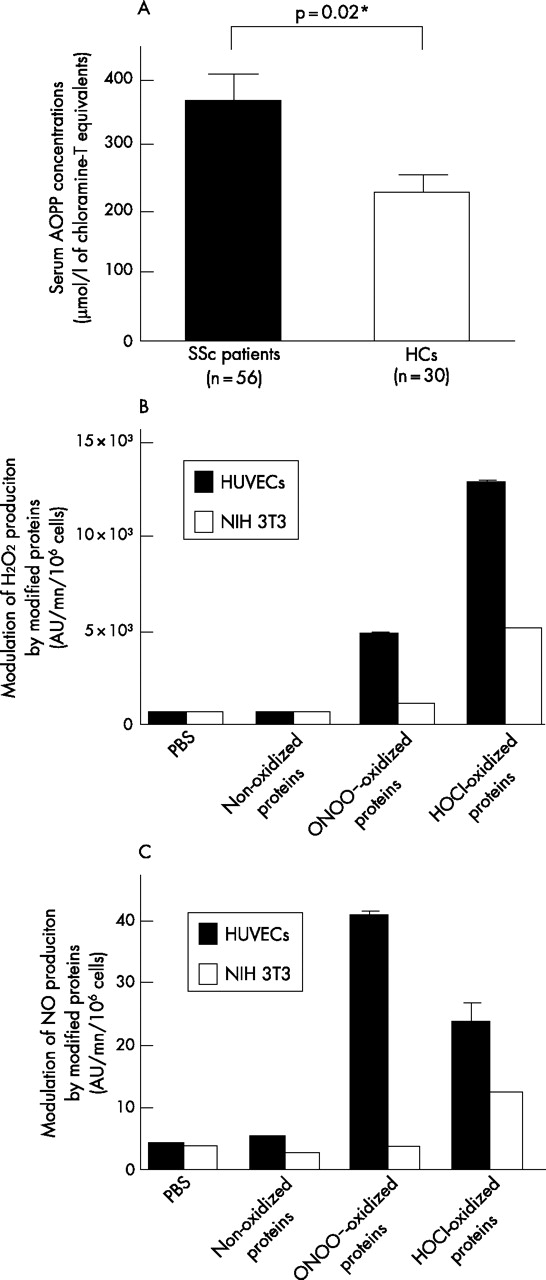
The serum concentration of AOPPs was higher in SSc patients than in healthy controls (p = 0.02; fig 3A) and was correlated with the induction of H2O2 production by HUVECs (r = 0.46, p<0.001), NIH 3T3 cells (r = 0.35, p = 0.01) and HEp-2 cells (r = 0.31, p = 0.02). HOCl-induced AOPP activated endothelial cells and, to a lesser extent, fibroblasts, whereas ONOO–-induced AOPP activated endothelial cells only. HOCl-AOPP induced more H2O2 than NO, whereas ONOO–-AOPP mainly induced NO (fig 3B and 3C). Furthermore, a dose-dependent proliferative response was observed with HOCl-AOPP and, to a lesser extent, with ONOO–-AOPP (data not shown).

Role of AOPPs in serum-induced ROS production. (A) Serum AOPP concentration in 56 SSc patients and in 30 healthy controls (HCs). Data are means ±SEM. (B) Production of hydrogen peroxide (H2O2) induced by HOCl-AOPP, peroxynitrites-AOPP or non-oxidized proteins by endothelial cells and fibroblasts. (C) Production of nitric oxide (NO) induced by HOCl-AOPP or peroxynitrites-AOPP or non-oxidized proteins in endothelial cells and fibroblasts. Data are means ±SEM. *Denotes statistical significance.
Disease-modifying agents modulate ROS generation and cellular proliferation induced by SSc sera
The ROS production and the fibroblast proliferation induced by the sera from seven patients (12.5%) who received immunosuppressive therapy at the time of serum collection was similar to those induced by SSc sera from non-treated patients. However, the sera from patients with immunosuppression abolished NO release by HUVECs (mean NO release: 10.14±2.3 vs 17.4±1.3; p = 0.049). Seven other patients (12.5%) received angiotensin-converting enzyme inhibitors. Their sera exerted the same effects as sera from SSc patients who did not receive this type of drug.
In vitro, no significant effect on H2O2 generation by HUVECs was observed in the presence of nifedipine, d-penicillamine or etanercept. Catalase, NAC and GSH decreased H2O2 levels by 23.4% (p = 0.006), 24.9% (p = 0.048) and 14.2% (p = 0.05), respectively (fig 4A). GSH increased HUVEC proliferation by 5% (p = 0.29) and NAC by 22.2% (p = 0.07), and catalase significantly improved the proliferation of HUVECs by 53% (p = 0.047) (data not shown).

Modulation of serum-induced ROS generation and cell proliferation by antioxidative drugs. (A) Modulation of hydrogen peroxide (H2O2) production by human umbilical venous endothelial cells (HUVECs). (B) Modulation of nitric oxide (NO) production by HUVECs. (C) Modulation of NIH 3T3 fibroblast proliferation. In A, B and C, HUVECs and fibroblasts were incubated with SSc sera and one of the following molecules: 10 μM nifedipine, 50 μM d-penicillamine, 50 μM bosentan, 10 μg etanercept, 1600 μM NAC, 400 μM reduced GSH or 10 U PEG-catalase. Results are expressed as percentages of ROS production or fibroblast proliferation versus untreated cells (100%). Error bars represent SEM.
Bosentan inhibited NO release by 32.7% (p<0.001) on HUVECs, whereas other molecules did not (fig 4B). GSH and catalase decreased SSc serum-induced production of H2O2 by NIH 3T3 cells by 19.8% (p = 0. 048) and by 27.8% (p = 0.044), respectively, whereas NAC did not (data not shown). GSH, d-penicillamine and 5FU inhibited fibroblast proliferation by 45.15% (p = 0.047), 71.8% (p<0.001) and 78.4% (p<0.001), respectively, whereas NAC alone did not (p = 0.15) (fig 4C). However, NAC potentiated the anti-proliferative activity of 5FU on fibroblasts by 10% (p = 0.02). Nifedipine, bosentan, etanercept and catalase had no or little effect on fibroblast proliferation (fig 4C).
Longitudinal study of in vitro ROS production by HUVECs and NIH 3T3 fibroblast proliferation
In four patients (patients A, B, C and D), clinical involvement worsened and immunosuppressive drug(s) was (were) given (fig 5A, 5B and 5C). AOPP concentrations and serum-induced H2O2 production decreased when the clinical status stabilised or improved. In patient E, no visceral involvement was detected in the course of the disease and serum properties were not modified when assayed longitudinally (fig 5C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Longitudinal study of ex vivo serum-induced ROS production by HUVECs, proliferation of NIH 3T3 fibroblast proliferation and serum AOPP concentrations in five patients. (A) Patient A had progressive restrictive syndrome (RS) with decreased XXX/vascular abnormalities (DLCO/VAs) and biopsy-proven inflammatory myositis. He received mycophenolate mofetyl and intravenous immunoglobulins. Test values from three successive sera obtained every 6 months are presented. (B) Patient B had diffuse systemic sclerosis (SSc) with progressive deterioration of the Rodnan’s score. Test values from three successive sera are presented. Azathioprin was given at the time of the collection of the first serum, then suspended and resumed at the time of collection of the third serum sample. (C) Patient C had a RS related to lung fibrosis. (D) Patient D had RS and digital ulcers. Both patients C and D had been receiving immunosuppressive drugs at the time of the second serum collection. Patient E had SSc without complication and received no immunosuppressant. AZA, azathioprin; CYC, cyclophosphamide; DLCO, carbon monoxide diffusing capacity; IVIg, intravenous immunoglobulins; MMF, mycophenolate mofetyl; TLC, total lung capacity.
DISCUSSION
This report highlights the involvement of the oxidative stress in SSc and describes new serum markers in order to assess the activity of the disease and predict the response to treatment.
A common feature of the SSc sera tested in this study is their ability to induce the production of H2O2, particularly by endothelial cells. As ROS can modulate cell activation and proliferation,17,18 and as fibroblasts and endothelial cells are selectively targeted in SSc, we tested the effects of SSc sera on the proliferation of murine and human fibroblasts and HUVECs. SSc sera exert a cell-specific proliferative effect, as the same sera that stimulate fibroblasts to proliferate, inhibit HUVEC proliferation and have no effect on HEp-2 cells. The difference observed between fibroblasts and endothelial cells could be related to the level of ROS produced, as a reverse correlation between the rate of proliferation and that of H2O2 production is observed in both types of cell. The high levels of H2O2 generated in endothelial cells probably reaches the threshold of toxicity and inhibits cell proliferation, whereas the proliferation of fibroblasts from SSc patients is directly correlated with intrinsic production of H2O2 as previously shown.6,19
We then observed that the sera from patients with a RS induce a higher production of H2O2 and a higher fibroblast proliferation than sera from other SSc patients. Those data are in agreement with the previous finding that the synthesis of collagen by fibroblasts from SSc patients is correlated with the oxidative stress.6,19
On the other hand, serum-induced NO release is associated with digital ischaemia or PAH, although beneficial vasodilating effects could have been expected from a high rate of NO production.20–24 The deleterious effect of NO might be related to its combination with other ROS to form highly toxic peroxynitrites, resulting in cell injuries and cell death.25,26
We next characterized the serum factors responsible for the oxidative stress. In our experience, the prevalence of auto-antibodies to endothelial cells and fibroblasts is that found usually in SSc,27–30 but those antibodies are not correlated with serum-induced ROS production or cell proliferation. Therefore, we turned to AOPPs, the concentration of which has been found to be elevated in SSc sera by us and others8,31 and that can trigger a respiratory burst in monocytes.16 In our experience, AOPPs activate endothelial cells and, to a lesser extent, fibroblasts to generate ROS. Furthermore, AOPPs generated by different oxidation patterns lead to the production of either NO or H2O2, suggesting that AOPPs may be involved in the generation of different types of ROS in SSc patients. However, the role of autoantibodies cannot be totally ruled out. For instance, anti-PDGF receptor antibodies32, and possibly environmental factors,4,33–35 could initiate a secondary self-maintained process3 that is linked to the production of AOPPs and ROS that perpetuate the disease.
We finally investigated whether drugs used in SSc and antioxidant molecules can modulate the pro-activating effects of SSc sera. NAC and GSH decrease the generation of H2O2 by HUVECs and the proliferation of fibroblasts following incubation with the majority of SSc sera. This is in accordance with the observation that NAC can decrease ROS production in SSc fibroblasts, inhibit their proliferation and collagen synthesis,6 and abrogate ONOO– synthesis by activated lung macrophages from SSc patients in vitro.36 In addition, we observed that NAC potentiates the anti-proliferative effect of 5FU, which is in agreement with the beneficial effect of NAC in association with immunosuppressive therapies observed in patients with lung idiopathic interstitial fibrosis.37
Bosentan, an antagonist of endothelin A and B receptors that exerts beneficial effects in SSc patients with pulmonary arterial hypertension,38–40 abrogates NO overproduction by HUVECs stimulated by sera from patients with vascular complications. Consequently, this drug could be preferentially used in SSc patients whose sera trigger NO release, prior to the development of severe vascular complications. Prospective therapeutic trials have to be performed in SSc patients to confirm these results.
In conclusion, our data underline a pivotal role of serum factors, such as ROS and AOPPs, in the pathogenesis of the different patterns observed in SSc. These serum markers could help in assessing the activity of SSc and selecting the most appropriate treatments according to the fibrotic or vascular pattern.
Acknowledgments
Amélie Servettaz received a grant from Actelion and the Association des Sclérodermiques de France. Philippe Guilpain received a grant from the Fondation pour la Recherche Médicale (FRM).
REFERENCES
Footnotes
-
Published Online First 15 March 2007
-
Competing interests: None declared